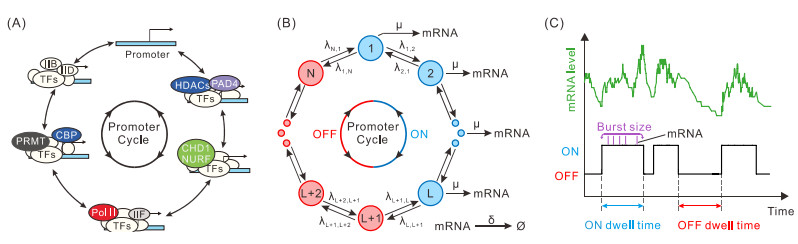
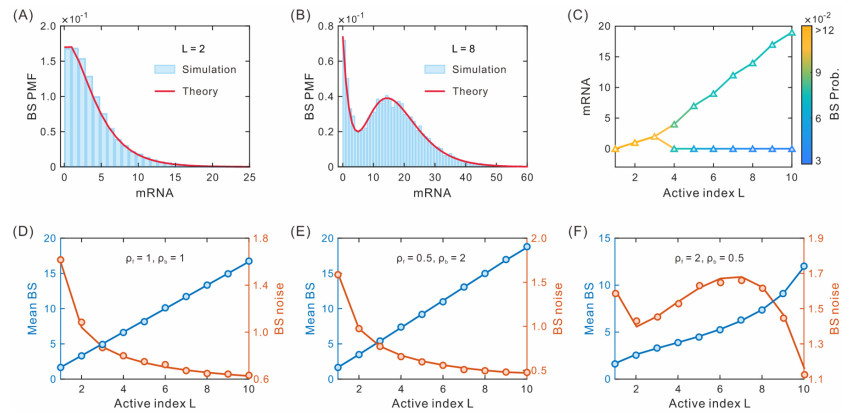
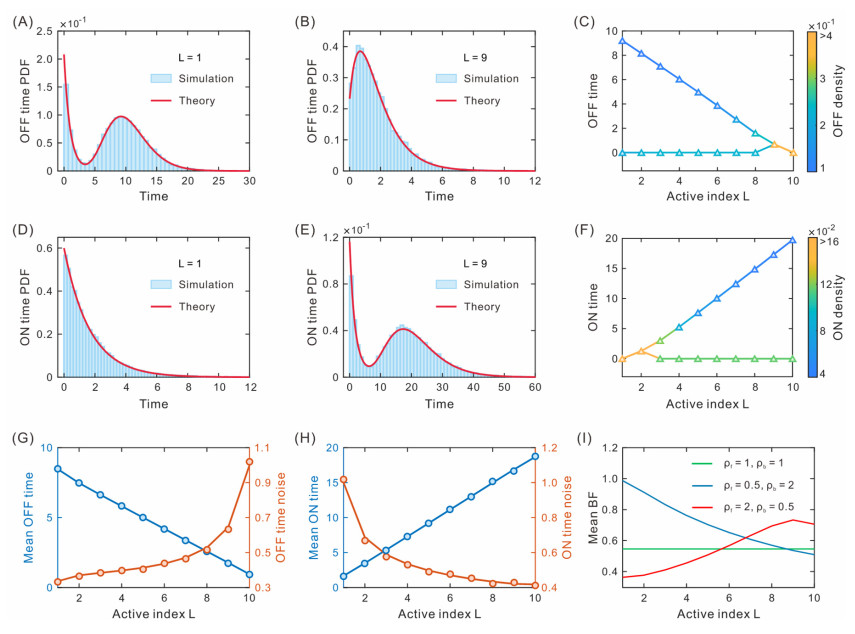
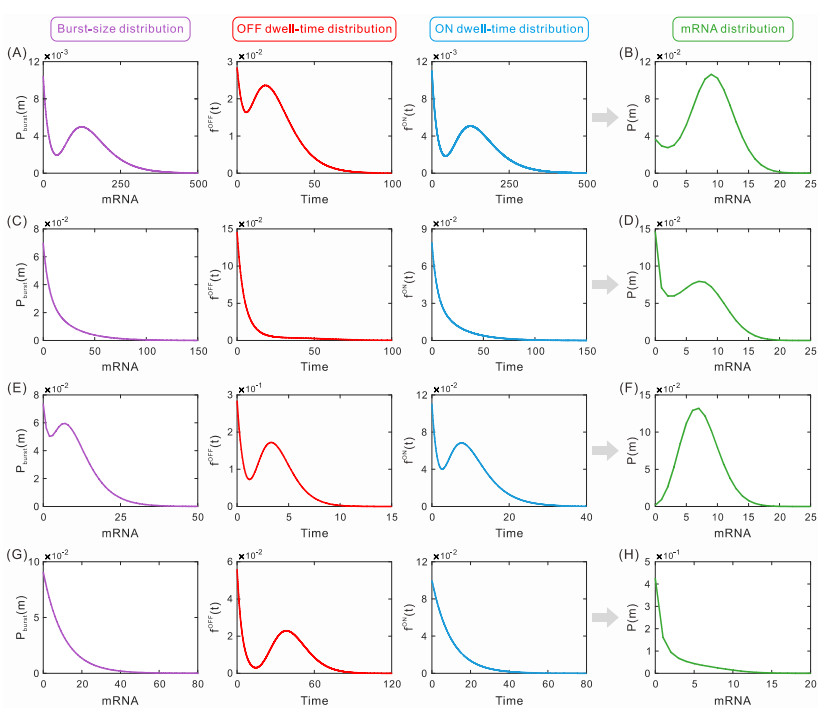
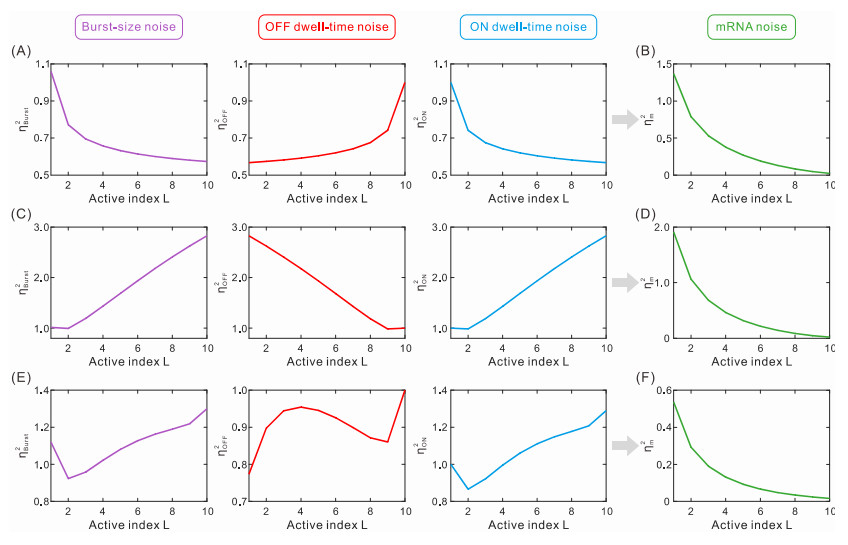
While transcription often occurs in a bursty manner, various possible regulations can lead to complex promoter patterns such as promoter cycles, giving rise to an important question: How do promoter kinetics shape transcriptional bursting kinetics? Here we introduce and analyze a general model of the promoter cycle consisting of multi-OFF states and multi-ON states, focusing on the effects of multi-ON mechanisms on transcriptional bursting kinetics. The derived analytical results indicate that burst size follows a mixed geometric distribution rather than a single geometric distribution assumed in previous studies, and ON and OFF times obey their own mixed exponential distributions. In addition, we find that the multi-ON mechanism can lead to bimodal burst-size distribution, antagonistic timing of ON and OFF, and diverse burst frequencies, each further contributing to cell-to-cell variability in the mRNA expression level. These results not only reveal essential features of transcriptional bursting kinetics patterns shaped by multi-state mechanisms but also can be used to the inferences of transcriptional bursting kinetics and promoter structure based on experimental data.
Citation: Xiyan Yang, Zihao Wang, Yahao Wu, Tianshou Zhou, Jiajun Zhang. Kinetic characteristics of transcriptional bursting in a complex gene model with cyclic promoter structure[J]. Mathematical Biosciences and Engineering, 2022, 19(4): 3313-3336. doi: 10.3934/mbe.2022153
While transcription often occurs in a bursty manner, various possible regulations can lead to complex promoter patterns such as promoter cycles, giving rise to an important question: How do promoter kinetics shape transcriptional bursting kinetics? Here we introduce and analyze a general model of the promoter cycle consisting of multi-OFF states and multi-ON states, focusing on the effects of multi-ON mechanisms on transcriptional bursting kinetics. The derived analytical results indicate that burst size follows a mixed geometric distribution rather than a single geometric distribution assumed in previous studies, and ON and OFF times obey their own mixed exponential distributions. In addition, we find that the multi-ON mechanism can lead to bimodal burst-size distribution, antagonistic timing of ON and OFF, and diverse burst frequencies, each further contributing to cell-to-cell variability in the mRNA expression level. These results not only reveal essential features of transcriptional bursting kinetics patterns shaped by multi-state mechanisms but also can be used to the inferences of transcriptional bursting kinetics and promoter structure based on experimental data.

| [1] |
I. Golding, J. Paulsson, S. M. Zawilski, E. C. Cox, Real-time kinetics of gene activity in individual bacteria, Cell, 123 (2005), 1025–1036. https://doi.org/10.1016/j.cell.2005.09.031 doi: 10.1016/j.cell.2005.09.031

|
| [2] |
A. Raj, C. S. Peskin, D. Tranchina, D. Y. Vargas, S. Tyagi, Stochastic mRNA synthesis in mammalian cells, PLoS Biol., 4 (2006), e309. https://doi.org/10.1371/journal.pbio.0040309 doi: 10.1371/journal.pbio.0040309
|
| [3] |
J. R. Chubb, T. Trcek, S. M. Shenoy, R. H. Singer, Transcriptional pulsing of a developmental gene, Curr. Biol., 16 (2006), 1018–1025. https://doi.org/10.1016/j.cub.2006.03.092 doi: 10.1016/j.cub.2006.03.092
|
| [4] |
L. So, A. Ghosh, C. Zong, L. A. Sepúlveda, R. Segev, I. Golding, General properties of transcriptional time series in Escherichia coli., Nat. Genet., 43 (2011), 554–560. https://doi.org/10.1038/ng.821 doi: 10.1038/ng.821
|
| [5] |
M. B. Elowitz, A. J. Levine, E. D. Siggia, P. S. Swain, Stochastic gene expression in a single cell, Science, 297 (2002), 1183–1186. https://doi.org/10.1126/science.1070919 doi: 10.1126/science.1070919
|
| [6] |
J. M. Raser, E. K. O'Shea, Control of stochasticity in eukaryotic gene expression, Science, 304 (2004), 1811–1184. https://doi.org/10.1126/science.1098641 doi: 10.1126/science.1098641
|
| [7] |
A. Becskei, B. B. Kaufmann, A. van Oudenaarden, Contributions of low molecule number and chromosomal positioning to stochastic gene expression, Nat. Genet., 37 (2005), 937–944. https://doi.org/10.1038/ng1616 doi: 10.1038/ng1616
|
| [8] |
G. Balázsi, A. van Oudenaarden, J. J. Collins, Cellular decision making and biological noise: from microbes to mammals, Cell, 144 (2011), 910–925. https://doi.org/10.1016/j.cell.2011.01.030 doi: 10.1016/j.cell.2011.01.030
|
| [9] |
A. N. Boettiger, M. Levine, Synchronous and stochastic patterns of gene activation in the Drosophila embryo, Science, 325 (2009), 471–473. https://doi.org/10.1126/science.1173976 doi: 10.1126/science.1173976
|
| [10] |
A. Raj, S. A. Rifkin, E. Andersen, A. van Oudenaarden, Variability in gene expression underlies incomplete penetrance, Nature, 463 (2010), 913–918. https://doi.org/10.1038/nature08781 doi: 10.1038/nature08781
|
| [11] |
A. Eldar, M. B. Elowitz, Functional roles for noise in genetic circuits, Nature, 467 (2010), 167–173. https://doi.org/10.1038/nature09326 doi: 10.1038/nature09326
|
| [12] |
P. J. Choi, L. Cai, K. Frieda, X. S. Xie, A stochastic single-molecule event triggers phenotype switching of a bacterial cell, Science, 322 (2008), 442–446. https://doi.org/10.1126/science.1161427 doi: 10.1126/science.1161427
|
| [13] |
M. Ptashne, A. Gann, Transcriptional activation by recruitment, Nature, 386 (1997), 569–577. https://doi.org/10.1038/386569a0 doi: 10.1038/386569a0
|
| [14] |
G. L. Hager, C. Elbi, T. A. Johnson, T. Voss, A. K. Nagaich, R. L. Schiltz, et al., Chromatin dynamics and the evolution of alternate promoter states, Chromosome Res., 14 (2006), 107–116. https://doi.org/10.1007/s10577-006-1030-0 doi: 10.1007/s10577-006-1030-0
|
| [15] |
C. R. Clapier, B. R. Cairns, The biology of chromatin remodeling complexes, Annu. Rev. Biochem., 78 (2009), 273–304. https://doi.org/10.1146/annurev.biochem.77.062706.153223 doi: 10.1146/annurev.biochem.77.062706.153223
|
| [16] |
T. J. Stasevich, J. G. McNally, Assembly of the transcription machinery: ordered and stable, random and dynamic, or both?, Chromosoma, 120 (2011), 533–545. https://doi.org/10.1007/s00412-011-0340-y doi: 10.1007/s00412-011-0340-y
|
| [17] |
D. R. Larson, What do expression dynamics tell us about the mechanism of transcription?, Curr. Opin. Genet. Dev., 21 (2011), 591–599. https://doi.org/10.1016/j.gde.2011.07.010 doi: 10.1016/j.gde.2011.07.010
|
| [18] |
D. M. Suter, N. Molina, D. Gatfield, K. Schneider, U. Schibler, F. Naef, Mammalian genes are transcribed with widely different bursting kinetics, Science, 332 (2011), 472–474. https://doi.org/10.1126/science.1198817 doi: 10.1126/science.1198817
|
| [19] |
C. V. Harper, B. Finkenstädt, D. J. Woodcock, S. Friedrichsen, S. Semprini, L. Ashall, et al., Dynamic analysis of stochastic transcription cycles, PLoS Biol., 9 (2011), e1000607. https://doi.org/10.1371/journal.pbio.1000607 doi: 10.1371/journal.pbio.1000607
|
| [20] |
M. Kandhavelu, A. Hӓkkinen, O. Yli-Harja, A. S. Ribeiro, Single-molecule dynamics of transcription of the lar promoter, Phys. Biol., 9 (2012), 026004. https://doi.org/10.1088/1478-3975/9/2/026004 doi: 10.1088/1478-3975/9/2/026004
|
| [21] |
W. J. Blake, G. Balázsi, M. A. Kohanski, F. J. Isaacs, K. F. Murphy, Y. Kuang, et al., Phenotypic consequences of promoter-mediated transcriptional noise, Mol. Cell., 24 (2006), 853–865. https://doi.org/10.1016/j.molcel.2006.11.003 doi: 10.1016/j.molcel.2006.11.003
|
| [22] |
S. L. Berger, The complex language of chromatin regulation during transcription, Nature, 447 (2007), 407–412. https://doi.org/10.1038/nature05915 doi: 10.1038/nature05915
|
| [23] |
N. J. Fuda, M. B. Ardehali, J. T. Lis, Defining mechanisms that regulate RNA polymerase II transcription in vivo, Nature, 461 (2009), 186–192. https://doi.org/10.1038/nature08449 doi: 10.1038/nature08449
|
| [24] |
G. Hornung, R. Bar-Ziv, D. Rosin, N. Tokuriki, D. S. Tawfik, M. Oren, et al., Noise-mean relationship in mutated promoters, Genome Res., 22 (2012), 2409–2417. https://doi.org/10.1101/gr.139378.112 doi: 10.1101/gr.139378.112
|
| [25] |
A. Halme, S. Bumgarner, C. Styles, G. R. Fink, Genetic and epigenetic regulation of the FLO gene family generates cell-surface variation in yeast, Cell, 116 (2004), 405–415. https://doi.org/10.1016/S0092-8674(04)00118-7 doi: 10.1016/S0092-8674(04)00118-7
|
| [26] |
L. M. Octavio, K. Gedeon, N. Maheshri, Epigenetic and conventional regulation is distributed among activators of FLO11 allowing tuning of population-level heterogeneity in its expression, PLoS. Genet., 5 (2009), e1000673. https://doi.org/10.1371/journal.pgen.1000673 doi: 10.1371/journal.pgen.1000673
|
| [27] |
G. Reid, R. Gallais, R. Métivier, Marking time: the dynamic role of chromatin and covalent modification in transcription, Int. J. Biochem. Cell Biol., 41 (2009), 155–163. https://doi.org/10.1016/j.biocel.2008.08.028 doi: 10.1016/j.biocel.2008.08.028
|
| [28] |
L. S. S. Dunham, H. Momiji, C. V. Harper, P. J. Downton, K. Hey, A. McNamara, et al., Asymmetry between activation and deactivation during a transcriptional pulse, Cell Syst., 5 (2017), 646–653. https://doi.org/10.1016/j.cels.2017.10.013 doi: 10.1016/j.cels.2017.10.013
|
| [29] |
L. A. Sepúlveda, H. Xu, J. Zhang, M. Wang, I. Golding, Measurement of gene regulation in individual cells reveals rapid switching between promoter states, Science, 351 (2016), 1218–1222. https://doi.org/10.1126/science.aad0635 doi: 10.1126/science.aad0635
|
| [30] |
G. Neuert, B. Munsky, R. Z. Tan, L. Teytelman, M. Khammash, A. van Oudenaarden, Systematic identification of signal-activated stochastic gene regulation, Science, 339 (2013), 584–587. https://doi.org/10.1126/science.1231456 doi: 10.1126/science.1231456
|
| [31] |
J. M. Pedraza, J. Paulsson, Effects of molecular memory and bursting on fluctuations in gene expression, Science, 319 (2008), 339–343. https://doi.org/10.1126/science.1144331 doi: 10.1126/science.1144331
|
| [32] |
T. Jia, R. V. Kulkarni, Intrinsic noise in stochastic models of gene expression with molecular memory and bursting, Phys. Rev. Lett., 106 (2011), 058102. https://doi.org/10.1103/PhysRevLett.106.058102 doi: 10.1103/PhysRevLett.106.058102
|
| [33] |
J. Zhang, T. Zhou, Markovian approaches to modeling intracellular reaction processes with molecular memory, Proc. Natl. Acad. Sci. USA, 116 (2019), 23542–23550. https://doi.org/10.1073/pnas.1913926116 doi: 10.1073/pnas.1913926116
|
| [34] |
X. Yang, Y. Chen, T. Zhou, J. Zhang, Exploring dissipative sources of non-Markovian biochemical reaction systems, Phys. Rev. E, 103 (2021), 052411. https://doi.org/10.1103/PhysRevE.103.052411 doi: 10.1103/PhysRevE.103.052411
|
| [35] |
L. B. Carey, D. van Dijk, P. M. A. Sloot, J. A. Kaandorp, E. Segal, Promoter sequence determines the relationship between expression level and noise, PLoS Biol., 11 (2013), e1001528. https://doi.org/10.1371/journal.pbio.1001528 doi: 10.1371/journal.pbio.1001528
|
| [36] |
Y. Wang, T. Ni, W. Wang, F. Liu, Gene transcription in bursting: a unified mode for realizing accuracy and stochasticity, Biol. Rev., 94 (2019), 248–258. https://doi.org/10.1111/brv.12452 doi: 10.1111/brv.12452
|
| [37] |
J. Peccoud, B. Ycart, Markovian modeling of gene-product synthesis, Theor. Pop. Biol., 48 (1995), 222–234. https://doi.org/10.1006/tpbi.1995.1027 doi: 10.1006/tpbi.1995.1027
|
| [38] |
V. Shahrezaei, P. S. Swain, Analytical distributions for stochastic gene expression, Proc. Nat. Acad. Sci. USA, 105 (2008), 17256–17261. https://doi.org/10.1073/pnas.0803850105 doi: 10.1073/pnas.0803850105
|
| [39] |
M. Dobrzyński, F. J. Bruggeman, Elongation dynamics shape bursty transcription and translation, Proc. Natl. Acad. Sci. USA, 106 (2009), 2583–2588. https://doi.org/10.1073/pnas.0803507106 doi: 10.1073/pnas.0803507106
|
| [40] |
R. D. Dar, B. S. Razooky, A. Singh, T. V. Trimeloni, J. M. McCollum, et al., Transcriptional burst frequency and burst size are equally modulated across the human genome, Proc. Nat. Acad. Sci. USA, 109 (2012), 17454–17459. https://doi.org/10.1073/pnas.1213530109 doi: 10.1073/pnas.1213530109
|
| [41] |
J. Zhang, T. Zhou, Stationary moments, distribution conjugation and phenotypic regions in stochastic gene transcription, Math. Biosci. Eng. 16 (2019), 6134–6166. https://doi.org/10.3934/mbe.2019307 doi: 10.3934/mbe.2019307
|
| [42] |
C. Shi, Y. Jia, T. Zhou, Queuing models of gene expression: analytical distributions and beyond, Biophys. J., 119 (2020), 1606–1616. https://doi.org/10.1016/j.bpj.2020.09.001 doi: 10.1016/j.bpj.2020.09.001
|
| [43] |
Z. Zhang, Q. Deng, Z. Wang, Y. Chen, T. Zhou, Exact results for queuing models of stochastic transcription with memory and crosstalk, Phys. Rev. E, 103 (2021), 062414. https://doi.org/10.1103/PhysRevE.103.062414 doi: 10.1103/PhysRevE.103.062414
|
| [44] |
A. Schwabe, K. N. Rybakova, F. J. Bruggeman, Transcription stochasticity of complex gene regulation models, Biophys. J., 103 (2012), 1152–1161. https://doi.org/10.1016/j.bpj.2012.07.011 doi: 10.1016/j.bpj.2012.07.011
|
| [45] |
N. Kumar, A. Singh, R. V. Kulkarni, Transcriptional bursting in gene expression: analytical results for general stochastic models, PLoS Comput. Biol., 11 (2015), e1004292. https://doi.org/10.1371/journal.pcbi.1004292 doi: 10.1371/journal.pcbi.1004292
|
| [46] |
J. Zhang, L. Chen, T. Zhou, Analytical distribution and tunability of noise in a model of promoter progress, Biophys. J., 102 (2012), 1247–1257. https://doi.org/10.1016/j.bpj.2012.02.001 doi: 10.1016/j.bpj.2012.02.001
|
| [47] |
T. Zhou, J. Zhang, Analytical results for a multistate gene model, SIAM J. Appl. Math. 72 (2012), 789-818. https://doi.org/10.1137/110852887 doi: 10.1137/110852887
|
| [48] |
J. Zhang, T. Zhou, Promoter-mediated transcription dynamics, Biophys. J., 106 (2014), 479–488. https://doi.org/10.1016/j.bpj.2013.12.011 doi: 10.1016/j.bpj.2013.12.011
|
| [49] |
S. Choubey, J. Kondev, A. Sanchez, Distribution of initiation times reveals mechanisms of transcriptional regulation in single cells, Biophys. J., 114 (2018), 2072–2082. https://doi.org/10.1016/j.bpj.2018.03.031 doi: 10.1016/j.bpj.2018.03.031
|
| [50] |
B. Zoller, D. Nicolas, N. Molina, F. Naef, Structure of silent transcription intervals and noise characteristics of mammalian genes, Mol. Syst. Biol., 11 (2015), 823. https://doi.org/10.15252/msb.20156257 doi: 10.15252/msb.20156257
|
| [51] |
K. N. Rybakova, F. J. Bruggeman, A. Tomaszewska, M. J. Moné, C. Carlberg, H. V. Westerhoff, Multiplex eukaryotic transcription (in) activation: timing, bursting and cycling of a ratchet clock mechanism, PLoS Comput. Biol., 11(2015), e1004236. https://doi.org/10.1371/journal.pcbi.1004236 doi: 10.1371/journal.pcbi.1004236
|
| [52] |
[52] B. J. Daigle, J. M. Soltani, L. R. Petzold, A. Singh, Inferring single-cell gene expression mechanisms using stochastic simulation, Bioinformatics, 31 (2015), 1428–1435. https://doi.org/10.1093/bioinformatics/btv007 doi: 10.1093/bioinformatics/btv007
|
| [53] |
T. L. To, N. Maheshri, Noise can induce bimodality in positive transcriptional feedback loops without bistability, Science, 327(2010), 1142. https://doi.org/10.1126/science.1178962 doi: 10.1126/science.1178962
|
| [54] |
P. Bokes, A. Borri, P. Palumbo, A. Singh, Mixture distributions in a stochastic gene expression model with delayed feedback: a WKB approximation approach, J. Math. Biol., 81 (2020) 343–367. https://doi.org/10.1007/s00285-020-01512-y doi: 10.1007/s00285-020-01512-y
|
| [55] |
P. Bokes, Postponing production exponentially enhances the molecular memory of a stochastic switch, Eur. J. Appl. Math., 2020 (2020), 1–18. https://doi.org/10.1017/S0956792520000479 doi: 10.1017/S0956792520000479
|
| [56] |
J. Zhang, Q. Nie, T. Zhou, A moment-convergence method for stochastic analysis of biochemical reaction networks, J. Chem. Phys., 144 (2016), 194109. https://doi.org/10.1063/1.4950767 doi: 10.1063/1.4950767
|
| [57] |
D. Gillespie, Exact stochastic simulation of coupled chemical reactions, J. Phys. Chem., 81 (1977), 2340–2361. https://doi.org/10.1021/j100540a008 doi: 10.1021/j100540a008
|
| [58] |
M. Kærn, T. C. Elston, W. J. Blake, J. J. Collins, Stochasticity in gene expression: from theories to phenotypes, Nat. Rev. Genet., 6 (2005), 451–464. https://doi.org/10.1038/nrg1615 doi: 10.1038/nrg1615
|
| [59] |
A. S. Hansen, E. K. O'Shea, Promoter decoding of transcription factor dynamics involves a trade-off between noise and control of gene expression, Mol. Syst. Biol., 9 (2013), 704. https://doi.org/10.1038/msb.2013.56 doi: 10.1038/msb.2013.56
|
| [60] |
Z. Cao, R. Grima, Analytical distributions for detailed models of stochastic gene expression in eukaryotic cells, Proc. Natl. Acad. Sci. USA, 117 (2020), 4682–4692. https://doi.org/10.1073/pnas.1910888117 doi: 10.1073/pnas.1910888117
|
| [61] |
Q. Wang, T. Zhou, Alternative-splicing-mediated gene expression, Phys. Rev. E, 89 (2014), 012713. https://doi.org/10.1103/PhysRevE.89.012713 doi: 10.1103/PhysRevE.89.012713
|
| [62] |
L. Fang, Z. Yuan, P. Liu, T. Zhou, Feedback-induced counterintuitive correlations of gene expression noise with bursting kinetics, Phys. Rev. E, 90 (2014), 052702. https://doi.org/10.1103/PhysRevE.90.052702 doi: 10.1103/PhysRevE.90.052702
|
Figures(5) / Tables(1)

Xiyan Yang, Zihao Wang, Yahao Wu, Tianshou Zhou, Jiajun Zhang. Kinetic characteristics of transcriptional bursting in a complex gene model with cyclic promoter structure[J]. Mathematical Biosciences and Engineering, 2022, 19(4): 3313-3336. doi: 10.3934/mbe.2022153







 DownLoad:
DownLoad: