Citation: Qiuying Li, Lifang Huang, Jianshe Yu. Modulation of first-passage time for bursty gene expression via random signals[J]. Mathematical Biosciences and Engineering, 2017, 14(5&6): 1261-1277. doi: 10.3934/mbe.2017065

| [1] | [ D. W. Adams,J. Errington, Bacterial cell division: Assembly, maintenance and disassembly of the Z ring, Nat. Rev. Microbiol., 7 (2009): 642-653. |
| [2] | [ W. J. Blake,M. KAErn,C. R. Cantor,J. J. Collins, Noise in eukaryotic gene expression, Nature, 422 (2003): 633-637. |
| [3] | [ H. Boeger,J. Griesenbeck,R. D. Kornberg, Nucleosome retention and the stochastic nature of promoter chromatin remodeling for transcription, Cell, 133 (2008): 716-726. |
| [4] | [ L. Cai,N. Friedman,X. S. Xie, Stochastic protein expression in individual cells at the single molecule level, Nature, 440 (2006): 358-362. |
| [5] | [ A. K. Chavali, V. C. Wong and K. Miller-Jensen, Distinct promoter activation mechanisms modulate noise-driven HIV gene expression, Sci. Rep., 5 (2015), 17661. |
| [6] | [ S. Chong,C. Y. Chen,H. Ge,X. S. Xie, Mechanism of transcriptional bursting in bacteria, Cell, 158 (2014): 314-326. |
| [7] | [ J. R. Chubb,T. B. Liverpool, Bursts and pulses: insights from single cell studies into transcriptional mechanisms, Curr. Opin. Genet. Dev., 20 (2010): 478-484. |
| [8] | [ R. D. Dar,N. N. Hosmane,M. R. Arkin, Screening for noise in gene expression identifies drug synergies, Science, 344 (2014): 1392-1396. |
| [9] | [ R. D. Dar, B. S. Razooky and A. Singh, etc., Transcriptional burst frequency and burst size are equally modulated across the human genome, Proc. Natl. Acad. Sci. USA., 109 (2012), 17454–17459. |
| [10] | [ J. J. Dennehy and I. N. Wang, Factors influencing lysis time stochasticity in bacteriophage $λ$, BMC Microbiol., 11 (2011), 174. |
| [11] | [ R. E. Dolmetsch,K. Xu,R. S. Lewis, Calcium oscillations increase the efficiency and specificity of gene expression, Nature, 392 (1998): 933-936. |
| [12] | [ V. Elgart, T. Jia, A. T. Fenley and R. Kulkarni, Connecting protein and mRNA burst distributions for stochastic models of gene expression, Phys. Biol., 8 (2011), 046001. |
| [13] | [ M. B. Elowitz,A. J. Levine,E. D. Siggia,P. S. Swaim, Stochastic gene expression in a single cell, Science, 297 (2002): 1183-1186. |
| [14] | [ N. Geva-zatorsky,E. Dekel,E. Batchelor,G. Lahav,U. Alon, Fourier analysis and systems identification of the p53 feedback loop, Proc. Natl. Acad. Sci. USA., 107 (2010): 13550-13555. |
| [15] | [ K. R. Ghusinga, C. A. Vargas-Garcia and A. Singh, A mechanistic stochastic framework for regulating bacterial cell division, Sci. Rep., 6 (2016), 30229. |
| [16] | [ T. Gregor,D. W. Tank,E. F. Wieschaus,W. Bialek, Probing the limits to positional information, Cell, 130 (2007): 153-164. |
| [17] | [ A. Gründing,M. D. Manson,R. Yong, Holins kill without warning, Proc. Natl. Acad. Sci. USA., 98 (2001): 9348-9352. |
| [18] | [ N. K. Grzimek,D. Dreis,S. Schmalz,M. J. Reddehase, Random, asynchronous, and asymmetric transcriptional activity of enhancer-flanking major immediate-early genes ie1/3 and ie2 during murine cytomegalovirus latency in the lungs, J. Virol, 75 (2001): 2692-2705. |
| [19] | [ T. Günther and A. Grundhoff, The epigenetic landscape of latent kaposi sarcoma-associated herpesvirus genomes, PLoS Pathog., 6 (2010), e1000935. |
| [20] | [ P. Guptasarma, Does replication-induced transcription regulate synthesis of the myriad low copy number proteins of Escherichia coli?, Bioessays, 17 (1995): 987-997. |
| [21] | [ P. Hänggi,P. Talkner,M. Borkovec, Reaction-rate theory: Fifty years after Kramers, Rev. Mod. Phys., 62 (1990): 251-341. |
| [22] | [ B Hu, D. A. Kessler, W. J. Rappel and H. Levine, Effects of Input noise on a simple biochemical switch, Phy. Rev. Lett., 107 (2011), 148101. |
| [23] | [ L. Huang, Z. Yuan, P. Liu and T. Zhou, Feedback-induced counterintuitive correlations of gene expression noise with bursting kinetics, Phys. Rev. E, 90 (2014), 052702. |
| [24] | [ P. J. Ingram, M. P. H. Stumpf and J. Stark, Nonidentifiability of the source of intrinsic noise in gene expression from single-burst data, PLoS Comput. Biol., 4 (2008), e1000192. |
| [25] | [ F. Jiao,M. Tang,J. Yu, Distribution profiles and their dynamic transition in stochastic gene transcription, J. Differential Equations, 254 (2013): 3307-3328. |
| [26] | [ I. G. Johnston, B. Gaal and R. P. Neves, et al., Mitochondrial variability as a source of extrinsic cellular noise, PloS Comput. Biol., 8 (2011), e1002416. |
| [27] | [ T. B. Kepler,T. C. Elston, Stochasticity in transcriptional regulation: Origins, consequences, and mathematical representations, Biophys. J., 81 (2001): 3116-3136. |
| [28] | [ T. K. Kim,R. Shiekhattar, Architectural and functional commonalities between enhancers and promoters, Cell, 162 (2015): 948-959. |
| [29] | [ D. M. Knipe and A. Cliffe, Chromatin control of herpes simplex virus lytic and latent infection, Nat. Rev. Microbiol., 6 (2008), 211-221, |
| [30] | [ J. Kuang,M. Tang,J. Yu, The mean and noise of protein numbers in stochastic gene expression, J. Math. Biol., 67 (2013): 261-291. |
| [31] | [ D. R. Larson, What do expression dynamics tell us about the mechanism of transcription?, Curr. Opin. Genet. Dev., 21 (2011): 591-599. |
| [32] | [ J. H. Levine,Y. Lin,M. B. Elowitz, Functional roles of pulsing in genetic circuits, Science, 342 (2013): 1193-1200. |
| [33] | [ W. Li,D. Notani,M. G. Rosenfeld, Enhancers as non-coding RNA transcription units: Recent insights and future perspectives, Nat. Rev. Genet., 17 (2016): 207-223. |
| [34] | [ Y. Li,M. Tang,J. Yu, Transcription dynamics of inducible genes modulated by negative regulations, Mathematical Medicine and Biology, 32 (2015): 115-136. |
| [35] | [ H. Maamar,A. Raj,D. Dubnau, Noise in gene expression determines cell fate in Bacillus subtilis, Science, 317 (2007): 526-529. |
| [36] | [ H. H. McAdams,A. Arkin, Stochastic mechanisms in gene expression, Proc. Natl. Acad. Sci. USA., 94 (1997): 814-819. |
| [37] | [ K. Miller-Jensen,S. S. Dey,D. V. Schaffer,A. P. Arkin, Varying virulence: Epigenetic control of expression noise and disease processes, Trends Biotechnol., 29 (2011): 517-525. |
| [38] | [ T. Morisaki,K. Lyon,K. F. DeLuca, Real-time quantification of single RNA translation dynamics in living cells, Science, 352 (2016): 1425-1429. |
| [39] | [ A. Ochab-marcinek,M. Tabaka, Bimodal gene expression in noncooperative regulatory systems, Proc. Natl. Acad. Sci. USA., 107 (2010): 22096-22101. |
| [40] | [ U. A. Orom,T. Derrien,M. Beringer,K. Gumireddy, Long noncoding RNAs with enhancer-like function in human cells, Cell, 143 (2010): 46-58. |
| [41] | [ M. Osella and M. C. Lagomarsino, Growthrate-dependent dynamics of a bacterial genetic oscillator, Phys. Rev. E, 87 (2013), 012726. |
| [42] | [ J. Pahle, A. K. Green, C. J. Dixon and U. Kummer, Information transfer in signaling pathways: A study using coupled simulated and experimental data, BMC Bioinformatics, 9 (2008), 139. |
| [43] | [ J. Paulsson,O. G. Berg,M. Ehrenberg, Stochastic focusing: fluctuation-enhanced sensitivity of intracellular regulation, Proc. Natl. Acad. Sci. USA., 97 (2000): 7148-7153. |
| [44] | [ J. M. Pedraza,J. Paulsson, Effects of molecular memory and bursting on fluctuations in gene expression, Science, 319 (2008): 339-343. |
| [45] | [ N. Petrenko,R. V. Chereji,M. N. McClean, Noise and interlocking signaling pathways promote distinct transcription factor dynamics in repond to different stresses, Mol. Biol. Cell, 24 (2013): 45-57. |
| [46] | [ A. Raj,A. V. Oudenaarden, Nature, nurture, or chance: Stochastic gene expression and its consequences, Cell, 135 (2008): 216-226. |
| [47] | [ J. M. Raser,E. K. O'Shea, Control of stochasticity in eukaryotic gene expression, Science, 304 (2004): 1811-1814. |
| [48] | [ L. Salmena,L. Poliseno,Y. Tay,L. Kats,P. P. Pandolfi, A ceRNA hypothesis: The Rosetta stone of a hidden RNA language, Cell, 146 (2011): 353-358. |
| [49] | [ A. Sanchez,S. Choubey,J. Kondev, Regulation of noise in gene expression, Annu. Rev. Biophys., 42 (2013): 469-491. |
| [50] | [ A. Sanchez,I. Golding, Genetic determinants and cellular constraints in noisy gene expression, Science, 342 (2013): 1188-1193. |
| [51] | [ V. Shahrezaei,P. S. Swain, Analytical distribution for stochastic gene expression, Proc. Natl. Acad. Sci. USA., 105 (2008): 17256-17261. |
| [52] | [ M. Shreshtha, A. Surendran and A. Ghosh, Estimation of mean first passage time for bursty gene expression, Phys. Biol., 13 (2016), 036004. |
| [53] | [ A. Singh and J. J. Dennehy, Stochastic holin expression can account for lysis time variation in the bacteriophage $λ$, J. R. Soc. Interface, 11 (2014), 20140140. |
| [54] | [ A. Singh and M. Soltani, Quantifying intrinsic and extrinsic variability in stochastic gene expression models, PLoS ONE, 8 (2013), e84301. |
| [55] | [ R. Skupsky, J. C. Burnett and J. E. Foley, et al., HIV promoter integration site primarily modulates transcriptional burst size rather than frequency, PLoS Comput. Biol., 6 (2010), e1000952. |
| [56] | [ M. Soltani, C. A. Vargas-Garcia, D. Antunes and A. Singh, Intercellular variability in protein levels from stochastic expression and noisy cell cycle processes, PLoS Comput. Biol. 12 (2016), e1004972. |
| [57] | [ G. M. Süel, R. P. Kulkarni and J. Dworkin, et al., Tunability and noise dependence in differentiation dynamics, Science, 315 (2007), 1137455. |
| [58] | [ Q. Sun,M. Tang,J. Yu, Temporal profile of gene transcription noise modulated by cross-talking signal transduction pathways, Bull. Math. Biol., 74 (2012): 375-398. |
| [59] | [ Q. Sun,M. Tang,J. Yu, Modulation of gene transcription noise by competing transcription factors, J. Math. Biol., 64 (2012): 469-494. |
| [60] | [ D. M. Suter,N. Molina,D. Gatfield,K. Schneider,U. Schibler,F. Naef, Mammalian genes are transcribed with widely different bursting kinetics, Science, 332 (2011): 472-474. |
| [61] | [ P. S. Swain,M. B. Elowitz,E. D. Siggia, Intrinsic and extrinsic contributions to stochasticity in gene expression, Proc. Natl. Acad. Sci. USA., 99 (2002): 12795-12800. |
| [62] | [ M. Thattai,O.A. Van, Intrinsic noise in gene regulatory networks, Proc. Natl. Acad. Sci. USA., 98 (2001): 8614-8619. |
| [63] | [ R. L. Thompson, C. M. Preston and N. M. Sawtell, De novo synthesis of vp16 coordinates the exit from hsv latency in vivo,PLoS Pathog., 5 (2009), e1000352. |
| [64] | [ F. Tostevin, R. W. De and P. R. Ten Wolde, Reliability of frequency and amplitude decoding in gene regulation, Phys. Rev. Lett., 108 (2012), 108104. |
| [65] | [ Z. Toth, D. T. Maglinte, S. H. Lee, H. R. Lee and L. Y. Wong, Epigenetic analysis of kshv latent and lytic genomes, PLoS Pathog. , 6 (2010), e1001013. |
| [66] | [ H. Wang, Z. Yuan, P. Liu and T. Zhou, Mechanisms of information decoding in a cascade system of gene expression, Phys. Rev. E, 93 (2016), 052411. |
| [67] | [ K. B. Wee,W. K. Yio,U. Surana,K. H. Chiam, Transcription factor oscillations induce differential gene expressions, Biophys. J., 102 (2012): 2413-2423. |
| [68] | [ S. L. Werner,D. Barken,A. Hoffmann, Stimulus specificity of gene expression programs determined by temporal control of IKK activity, Science, 309 (2005): 1857-1861. |
| [69] | [ J. Yu,Q. Sun,M. Tang, The nonlinear dynamics and fluctuations of mRNA levels in cross-talking pathway activated transcription, Journal Theoretical Biology, 363 (2014): 223-234. |
| [70] | [ J. Yu,J. Xiao,X. Ren,K. Lao,X. S. Xie, Probing gene expression in live cells, one protein molecule at a time, Science, 311 (2006): 1600-1603. |
| [71] | [ J. Zhang,L. Chen,T. Zhou, Analytical distribution and tunability of noise in a model of promoter progress, Biophys. J., 102 (2012): 1247-1257. |
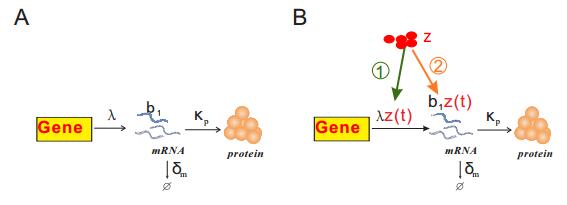
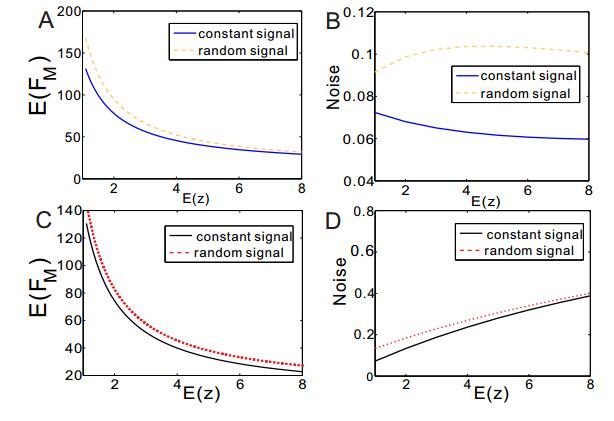

Figures(3)

Qiuying Li, Lifang Huang, Jianshe Yu. Modulation of first-passage time for bursty gene expression via random signals[J]. Mathematical Biosciences and Engineering, 2017, 14(5&6): 1261-1277. doi: 10.3934/mbe.2017065







 DownLoad:
DownLoad: