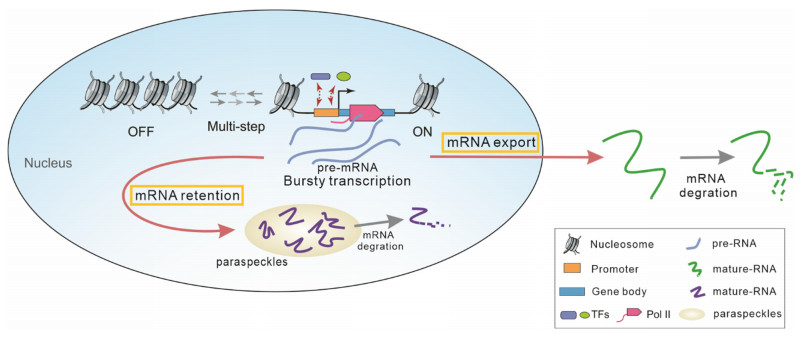
Transcription involves gene activation, nuclear RNA export (NRE) and RNA nuclear retention (RNR). All these processes are multistep and biochemical. A multistep reaction process can create memories between reaction events, leading to non-Markovian kinetics. This raises an unsolved issue: how does molecular memory affect stochastic transcription in the case that NRE and RNR are simultaneously considered? To address this issue, we analyze a non-Markov model, which considers multistep activation, multistep NRE and multistep RNR can interpret many experimental phenomena. In order to solve this model, we introduce an effective transition rate for each reaction. These effective transition rates, which explicitly decode the effect of molecular memory, can transform the original non-Markov issue into an equivalent Markov one. Based on this technique, we derive analytical results, showing that molecular memory can significantly affect the nuclear and cytoplasmic mRNA mean and noise. In addition to the results providing insights into the role of molecular memory in gene expression, our modeling and analysis provide a paradigm for studying more complex stochastic transcription processes.
Citation: Qiqi Deng, Aimin Chen, Huahai Qiu, Tianshou Zhou. Analysis of a non-Markov transcription model with nuclear RNA export and RNA nuclear retention[J]. Mathematical Biosciences and Engineering, 2022, 19(8): 8426-8451. doi: 10.3934/mbe.2022392
Transcription involves gene activation, nuclear RNA export (NRE) and RNA nuclear retention (RNR). All these processes are multistep and biochemical. A multistep reaction process can create memories between reaction events, leading to non-Markovian kinetics. This raises an unsolved issue: how does molecular memory affect stochastic transcription in the case that NRE and RNR are simultaneously considered? To address this issue, we analyze a non-Markov model, which considers multistep activation, multistep NRE and multistep RNR can interpret many experimental phenomena. In order to solve this model, we introduce an effective transition rate for each reaction. These effective transition rates, which explicitly decode the effect of molecular memory, can transform the original non-Markov issue into an equivalent Markov one. Based on this technique, we derive analytical results, showing that molecular memory can significantly affect the nuclear and cytoplasmic mRNA mean and noise. In addition to the results providing insights into the role of molecular memory in gene expression, our modeling and analysis provide a paradigm for studying more complex stochastic transcription processes.

| [1] |
J. Peccoud, B. Ycart, Markovian modelling of gene product synthesis, Theor. Popul. Biol., 48 (1995), 222-234. https://doi.org/10.1006/tpbi.1995.1027 doi: 10.1006/tpbi.1995.1027

|
| [2] |
V. Shahrezaei, P. S. Swain, Analytical distributions for stochastic gene expression, Proc. Natl. Acad. Sci. U.S.A., 105 (2008), 17256-17261. https://doi.org/10.1073/pnas.0803850105 doi: 10.1073/pnas.0803850105
|
| [3] |
J. Paulsson, Models of stochastic gene expression, Phys. Life Rev., 2 (2005), 157-175. https://doi.org/10.1016/j.plrev.2005.03.003 doi: 10.1016/j.plrev.2005.03.003
|
| [4] | T. B. Kepler, T. C. Elston, Stochasticity in transcriptional regulation: Origins, consequences, and mathematical representations, Biophys. J., 81 (2001), 3116-3036. https://doi.org/10.1016/S0006-3495(01)75949-8 |
| [5] |
N. Friedman, L. Cai, X. S. Xie, Linking stochastic dynamics to population-distribution: An analytical framework of gene expression, Phys. Rev. Lett., 97 (2006), 168302. https://doi.org/10.1103/PhysRevLett.97.168302 doi: 10.1103/PhysRevLett.97.168302
|
| [6] |
J. J. Zhang, T. S. Zhou, Promoter architecture-mediated transcriptional dynamics, Biophys. J., 106 (2014), 479-488. https://doi.org/10.1016/j.bpj.2013.12.011 doi: 10.1016/j.bpj.2013.12.011
|
| [7] |
J. J. Zhang, T. S. Zhou, Markovian approaches to modeling intracellular reaction processes with molecular memory, Proc. Natl. Acad. Sci. U.S.A., 116 (2019), 23542-23550. https://doi.org/10.1073/pnas.1913926116 doi: 10.1073/pnas.1913926116
|
| [8] |
T. Jia, R. V. Kulkarni, Intrinsic noise in stochastic models of gene expression with molecular memory and bursting, Phys. Rev. Lett., 106 (2011), 058102. https://doi.org/10.1103/PhysRevLett.106.058102 doi: 10.1103/PhysRevLett.106.058102
|
| [9] |
S. Choubey, Nascent RNA kinetics: Transient and steady state behavior of models of transcription, Phys. Rev. E., 97 (2018), 022402. https://doi.org/10.1103/PhysRevE.97.022402 doi: 10.1103/PhysRevE.97.022402
|
| [10] |
B. Zoller, S. C. Little, T. Gregor, Diverse spatial expression patterns emerge from unified kinetics of transcriptional bursting, Cell, 175 (2018), 835-847. https://doi.org/10.1016/j.cell.2018.09.056 doi: 10.1016/j.cell.2018.09.056
|
| [11] |
M. Z. Ali1, S. Choubey, Decoding the grammar of transcriptional regulation from RNA polymerase measurements: models and their applications, Phys. Biol., 16 (2019), 061001. https://doi.org/10.1088/1478-3975/ab45bf doi: 10.1088/1478-3975/ab45bf
|
| [12] |
T. B. Kallehauge, M. C. Robert, E. Bertrand, T. H. Jensen, Nuclear retention prevents premature cytoplasmic appearance of mRNA, Mol. Cell., 48 (2012), 145-152. https://doi.org/10.1016/j.molcel.2012.07.022 doi: 10.1016/j.molcel.2012.07.022
|
| [13] |
C. Marchive, F. Roudier, L. Castaings, V. Brehaut, E. Blondet, V. Colot, et al., Nuclear retention of the transcription factor NLP7 orchestrates the early response to nitrate in plants, Nat. Commun., 4 (2013), 1713. https://doi.org/10.1038/ncomms2650 doi: 10.1038/ncomms2650
|
| [14] |
Q. L. Zheng, J. Hou, Y. Zhou, Z. Y. Li, X. T. Cao, The RNA helicase DDX46 inhibits innate immunity by entrapping m6A-demethylated antiviral transcripts in the nucleus, Nat. Immunol., 18 (2017), 1094-1103. https://doi.org/10.1038/ni.3830 doi: 10.1038/ni.3830
|
| [15] |
A. Singh, P. Bokes, Consequences of mRNA transport on stochastic variability in protein levels, Biophys. J., 103 (2012), 1087-1096. https://doi.org/10.1016/j.bpj.2012.07.015 doi: 10.1016/j.bpj.2012.07.015
|
| [16] |
A. Mor, S. Suliman, R. Ben-Yishay, S. Yunger, Y. Brody, Y. Shav-Tal, Dynamics of single mRNP nucleocytoplasmic transport and export through the nuclear pore in living cells, Nat. Cell. Biol., 12 (2010), 543-552. https://doi.org/10.1038/ncb2056 doi: 10.1038/ncb2056
|
| [17] |
M. Oeffinger, D. Zenklusen, To the pore and through the pore: a story of mRNA export kinetics, Biochim. Biophys. Acta., 1819 (2012), 494-506. https://doi.org/10.1016/j.bbagrm.2012.02.011 doi: 10.1016/j.bbagrm.2012.02.011
|
| [18] |
V. O. Wickramasinghe, R. A. Laskey, Control of mammalian gene expression by selective mRNA export, Nat. Rev. Mol. Cell. Biol., 16 (2015), 431-442. https://doi.org/10.1038/nrm4010 doi: 10.1038/nrm4010
|
| [19] |
T. Chen, B. van Steensel, Comprehensive analysis of nucleocytoplasmic dynamics of mRNA in Drosophila cells, PLoS Genet., 13 (2017), e1006929. https://doi.org/10.1371/journal.pgen.1006929 doi: 10.1371/journal.pgen.1006929
|
| [20] |
N. Battich, T. Stoeger, L. Pelkmans, Control of transcript variability in single mammalian cells, Cell, 163 (2015), 1596-1610. https://doi.org/10.1016/j.cell.2015.11.018 doi: 10.1016/j.cell.2015.11.018
|
| [21] | G. L. Hager, C. Elbi, T. A. Johnson, T. Voss, A. K. Nagaich, R. L. Schiltz et al., Chromatin dynamics and the evolution of alternate promoter states. Chromosome Res., 14 (2006), 107-116. https://doi.org/10.1007/s10577-006-1030-0 |
| [22] |
R. Metivier, G. Reid, F. Gannon, Transcription in four dimensions: nuclear receptor-directed Transcription in four dimensions, EMBO Rep., 7 (2006), 161-167. https://doi.org/10.1038/sj.embor.7400626 doi: 10.1038/sj.embor.7400626
|
| [23] |
C. V. Harper, B. Finkenstädt, M. R. White, Dynamic analysis of stochastic transcription cycles, PLoS Biol., 9 (2011), e1000607. https://doi.org/10.1371/journal.pbio.1000607 doi: 10.1371/journal.pbio.1000607
|
| [24] |
Q. L. Wang, T. S. Zhou, Dynamical analysis of mCAT2 gene models with CTN-RNA nuclear retention, Phys. Biol., 12 (2015), 016010. https://doi.org/10.1088/1478-3975/12/1/016010 doi: 10.1088/1478-3975/12/1/016010
|
| [25] |
H. H. Qiu, B. G. Zhang, T. S. Zhou, Analytical results for a generalized model of bursty gene expression with molecular memory, Phys. Rev. E, 100 (2019), 012128. https://doi.org/10.1103/PhysRevE.100.012128 doi: 10.1103/PhysRevE.100.012128
|
| [26] |
K. B. Halpern, I. Caspi, D. Lemze, M. Levy, S. Landen, E. Elinav, et al., Nuclear retention of mRNA in mammalian tissues, Cell. Rep., 13 (2015), 2653-2662. https://doi.org/10.1016/j.celrep.2015.11.036 doi: 10.1016/j.celrep.2015.11.036
|
| [27] |
F. Alber, S. Dokudovskaya, L. M. Veenhoff, W. H. Zhang, J. Kipper, D. Devos, et al., The molecular architecture of the nuclear pore complex, Nature, 450 (2007), 695-701. https://doi.org/10.1038/nature06405 doi: 10.1038/nature06405
|
| [28] |
S. Choubey, J. Kondev, A. Sanchez, Deciphering transcriptional dynamics in vivo by counting nascent RNA molecules, PLoS Comput. Biol., 11 (2015), e1004345. https://doi.org/10.1371/journal.pcbi.1004345 doi: 10.1371/journal.pcbi.1004345
|
| [29] |
H. Xu, S. O. Skinner, A. M. Sokac, I. Golding, Stochastic kinetics of Nascent RNA, Phys. Rev. Lett, 117 (2016), 128101. https://doi.org/10.1103/PhysRevLett.117.128101 doi: 10.1103/PhysRevLett.117.128101
|
| [30] |
D. L. Spector, Nuclear domains, J. Cell. Sci., 114 (2001), 2891-2893. https://doi.org/10.1242/jcs.114.16.2891 doi: 10.1242/jcs.114.16.2891
|
| [31] |
A. H. Fox, Y. W. Lam, A. K. L. Leung, C. E. Lyon, J. Andersen, M. Mann, et al., Paraspeckles: A novel nuclear domain, Curr. Biol., 12 (2002), 13-25. https://doi.org/10.1016/S0960-9822(01)00632-7 doi: 10.1016/S0960-9822(01)00632-7
|
| [32] |
A. I. Lamond, D. L. Spector, Nuclear speckles: A model for nuclear organelles, Nat. Rev. Mol. Cell. Biol., 4 (2003), 605-612. https://doi.org/10.1038/nrm1172 doi: 10.1038/nrm1172
|
| [33] |
T. Aquino, M. Dentz, Chemical continuous time random walks, Phys. Rev. Lett, 119 (2017), 230601. https://doi.org/10.1103/PhysRevLett.119.230601 doi: 10.1103/PhysRevLett.119.230601
|
| [34] |
B. C. Arnold, Majorization: Here, there and everywhere, Statist. Sci., 22 (2007), 407-413. https://doi.org/10.1214/0883423060000000097 doi: 10.1214/0883423060000000097
|
| [35] |
A. David, S. Larry, The least variable phase type distribution is Erlang, Stoch. Models, 3 (1987), 467-473. https://doi.org/10.1080/15326348708807067 doi: 10.1080/15326348708807067
|
| [36] |
D. T. Gillespie, A general method for numerically simulating the stochastic time evolution of coupled chemical reactions, J. Comput. Phys, 22 (1976), 403-434. https://doi.org/10.1016/0021-9991(76)90041-3 doi: 10.1016/0021-9991(76)90041-3
|
| [37] |
I. Brouwer, T. L. Lenstra, Visualizing transcription: Key to understanding gene expression dynamics, Curr. Opin. Chem. Biol., 51 (2019), 122-129. https://doi.org/10.1016/j.cbpa.2019.05.031 doi: 10.1016/j.cbpa.2019.05.031
|
| [38] |
E. M. Wissink, A. Vihervaara, N. D. Tippens, J. T. Lis, Nascent RNA analyses: Tracking transcription and its regulation, Nat. Rev. Genet., 20 (2019), 705-723. https://doi.org/10.1038/s41576-019-0159-6 doi: 10.1038/s41576-019-0159-6
|
| [39] |
D. Grün, L. Kester, A. van Oudenaarden, Validation of noise models for single-cell transcriptomics, Nat. Methods., 11 (2014), 637. https://doi.org/10.1038/nmeth.2930 doi: 10.1038/nmeth.2930
|
| [40] |
D. R. Larson, R. H. Singer, D. Zenklusen, A single molecule view of gene expression, Cell. Biol., 19 (2009), 630-637. https://doi.org/10.1016/j.tcb.2009.08.008 doi: 10.1016/j.tcb.2009.08.008
|
| [41] |
A. E. Hage, S. L. French, A. L. Beyer, D. Tollervey, Loss of Topoisomerase I leads to R-loop-mediated transcriptional blocks during ribosomal RNA synthesis, Genes. Dev., 24 (2010), 1546-1558. https://doi.org/10.1101/gad.573310 doi: 10.1101/gad.573310
|
| [42] |
D. Grunwald, R. H. Singer, M. Rout, Nuclear export dynamics of RNA-protein complexes, Nature, 475 (2011), 333-341. https://doi.org/10.1038/nature10318 doi: 10.1038/nature10318
|
| [43] |
R. Benyishay, A. J. Ashkenazy, Y. Shav-Tal, Dynamic encounters of genes and transcripts with the nuclear pore, Trends. Genet., 32 (2016), 419-431. https://doi.org/10.1016/j.tig.2016.04.003 doi: 10.1016/j.tig.2016.04.003
|
| [44] |
Q. L. Wang, T. S. Zhou, Alternative-splicing-mediated gene expression, Phys. Rev. E., 89 (2014), 012713. https://doi.org/10.1103/PhysRevE.89.012713 doi: 10.1103/PhysRevE.89.012713
|
| [45] |
S. J. Gandhi, D. Zenklusen, T. Lionnet, R. H. Singer, Transcription of functionally related constitutive genes is not coordinated, Nat. Struct. Mol. Biol., 18 (2011), 27-34. https://doi.org/10.1038/nsmb.1934 doi: 10.1038/nsmb.1934
|
| [46] |
T. Stoeger, N. Battich, L. Pelkmans, Passive noise filtering by cellular compartmentalization, Cell, 164 (2016), 1151-1161. https://doi.org/10.1016/j.cell.2016.02.005 doi: 10.1016/j.cell.2016.02.005
|
| [47] | N. G. van Kampen, Stochastic processes in physics and chemistry, North-Holland, Amsterdam, 2007. https://doi.org/10.1016/B978-044452965-7/50006-4 |
| [48] |
J. Elf, M. Ehrenberg, Fast evaluation of fluctuations in biochemical networks with the linear noise approximation, Gen. Res, 13 (2003), 2475-2484. https://doi.org/10.1101/gr.1196503 doi: 10.1101/gr.1196503
|





Figures(6)

Qiqi Deng, Aimin Chen, Huahai Qiu, Tianshou Zhou. Analysis of a non-Markov transcription model with nuclear RNA export and RNA nuclear retention[J]. Mathematical Biosciences and Engineering, 2022, 19(8): 8426-8451. doi: 10.3934/mbe.2022392







 DownLoad:
DownLoad: