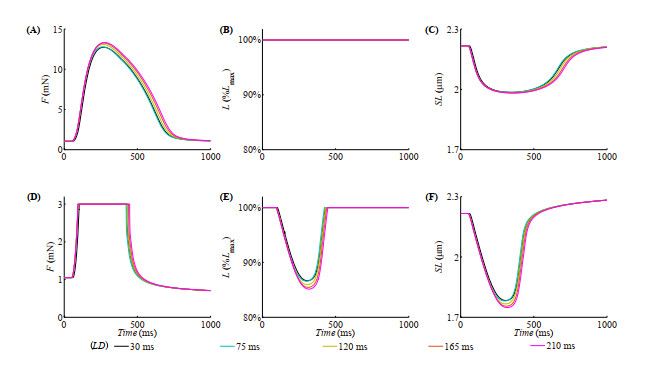
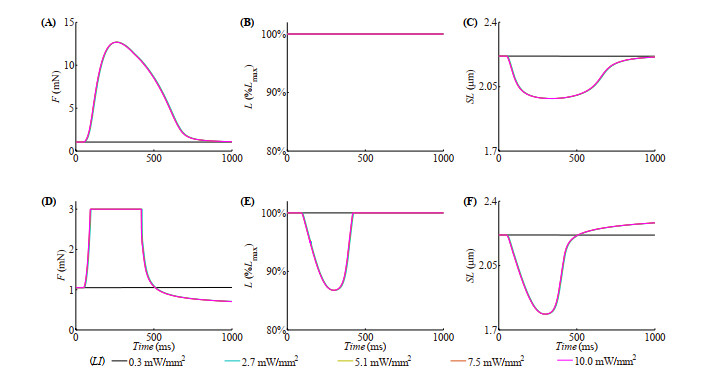
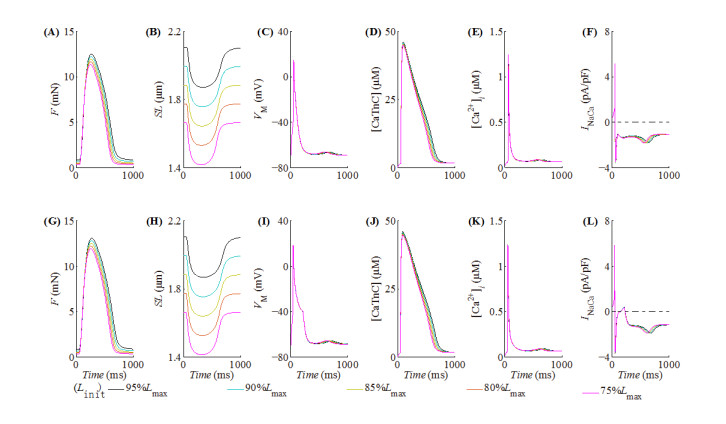
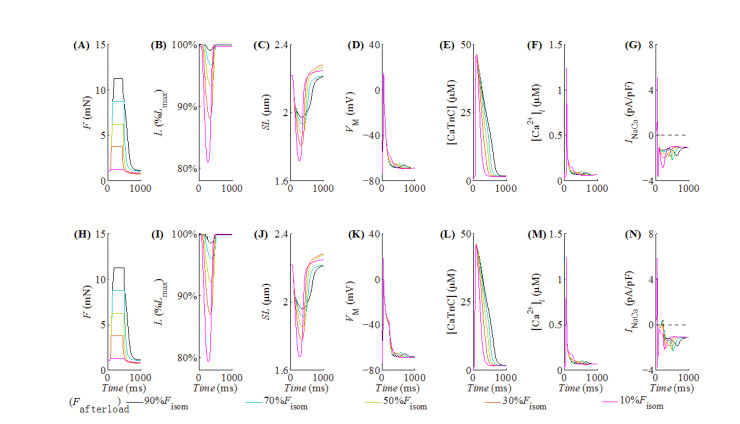
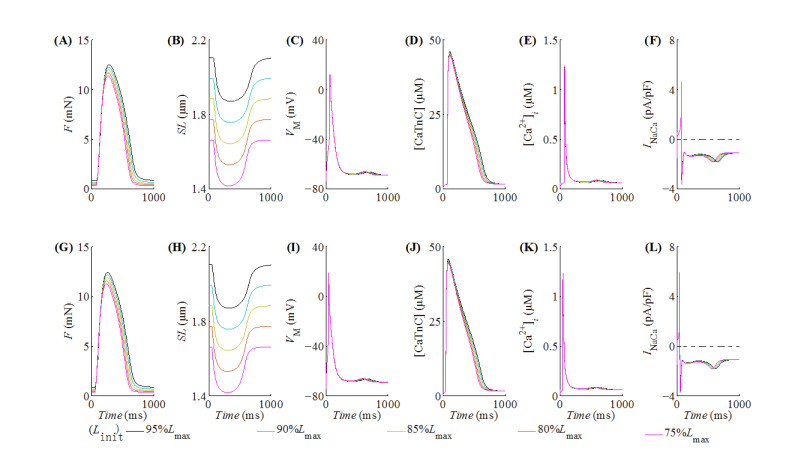
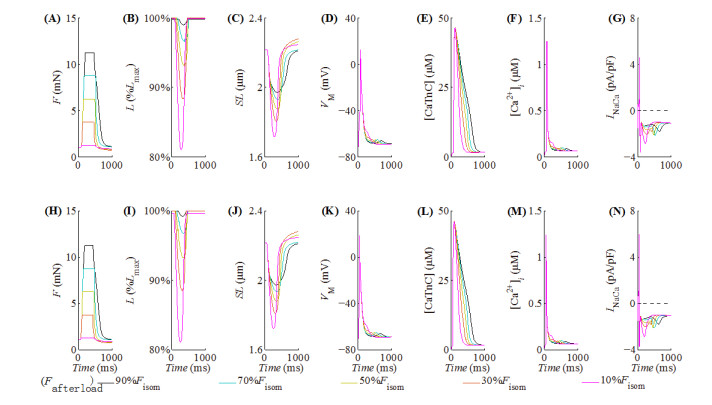
With the help of the conventional electrical method and the growing optogenetic technology, cardiac fibroblasts (Fbs) have been verified to couple electrically with working myocytes and bring electrophysiological remodeling changes in them. The intrinsic properties of cardiac functional autoregulation represented by excitation-contraction coupling (ECC) and mechano-electric feedback (MEF) have also been extensively studied. However, the roles of optogenetic stimulation on the characteristics of ECC and MEF in cardiomyocytes (CMs) coupled with Fbs have been barely investigated. In this study, we proposed a combined model composed of three modules to explore these influences. Simulation results showed that (1) during ECC, an increased light duration (LD) strengthened the inflow of ChR2 current and prolonged action potential duration (APD), and extended durations of twitch and internal sarcomere deformation through the decreased dissociation of calcium with troponin C (CaTnC) complexes and the prolonged duration of Xb attachment-detachment; (2) during MEF, an increased LD was followed by a longer muscle twitch and deformation, and led to APD prolongation through the inward ChR2 current and its inward rectification kinetics, which far outweighed the effects of the delaying dissociation of CaTnC complexes and the prolonged reverse mode of Na+-Ca2+ exchange on AP shortening; (3) due to the ChR2 current's rectification feature, enhancing the light irradiance (LI) brought slight variations in peak or valley values of electrophysiological and mechanical parameters while did not change durations of AP and twitch and muscle deformation in both ECC and MEF. In conclusion, the inward ChR2 current and its inward rectification feature were found to affect significantly the durations of AP and twitch in both ECC and MEF. The roles of optogenetic actuation on both ECC and MEF should be considered in future cardiac computational optogenetics at the tissue and organ scale.
Citation: Heqing Zhan, Zefeng Wang, Jialun Lin, Yuanbo Yu, Ling Xia. Optogenetic actuation in ChR2-transduced fibroblasts alter excitation-contraction coupling and mechano-electric feedback in coupled cardiomyocytes: a computational modeling study[J]. Mathematical Biosciences and Engineering, 2021, 18(6): 8354-8373. doi: 10.3934/mbe.2021414
With the help of the conventional electrical method and the growing optogenetic technology, cardiac fibroblasts (Fbs) have been verified to couple electrically with working myocytes and bring electrophysiological remodeling changes in them. The intrinsic properties of cardiac functional autoregulation represented by excitation-contraction coupling (ECC) and mechano-electric feedback (MEF) have also been extensively studied. However, the roles of optogenetic stimulation on the characteristics of ECC and MEF in cardiomyocytes (CMs) coupled with Fbs have been barely investigated. In this study, we proposed a combined model composed of three modules to explore these influences. Simulation results showed that (1) during ECC, an increased light duration (LD) strengthened the inflow of ChR2 current and prolonged action potential duration (APD), and extended durations of twitch and internal sarcomere deformation through the decreased dissociation of calcium with troponin C (CaTnC) complexes and the prolonged duration of Xb attachment-detachment; (2) during MEF, an increased LD was followed by a longer muscle twitch and deformation, and led to APD prolongation through the inward ChR2 current and its inward rectification kinetics, which far outweighed the effects of the delaying dissociation of CaTnC complexes and the prolonged reverse mode of Na+-Ca2+ exchange on AP shortening; (3) due to the ChR2 current's rectification feature, enhancing the light irradiance (LI) brought slight variations in peak or valley values of electrophysiological and mechanical parameters while did not change durations of AP and twitch and muscle deformation in both ECC and MEF. In conclusion, the inward ChR2 current and its inward rectification feature were found to affect significantly the durations of AP and twitch in both ECC and MEF. The roles of optogenetic actuation on both ECC and MEF should be considered in future cardiac computational optogenetics at the tissue and organ scale.

| [1] |
A. R. Pinto, A. Ilinykh, M. J. Ivey, J. T. Kuwabara, M. L. D'Antoni, R. Debuque, et al., Revisiting Cardiac Cellular Composition, Circ. Res., 118 (2016), 400-409. doi: 10.1161/CIRCRESAHA.115.307778

|
| [2] | J. Mayourian, D. K. Ceholski, D. M. Gonzalez, T. J. Cashman, S. Sahoo, R. J. Hajjar, et al., Physiologic, Pathologic, and Therapeutic Paracrine Modulation of Cardiac Excitation-Contraction Coupling, Circ. Res., 122 (2018), 167-183. |
| [3] |
D. M. Pedrotty, R. Y. Klinger, R. D. Kirkton, N. Bursac, Cardiac fibroblast paracrine factors alter impulse conduction and ion channel expression of neonatal rat cardiomyocytes, Cardiovasc. Res., 83 (2009), 688-697. doi: 10.1093/cvr/cvp164
|
| [4] | K. Kaur, M. Zarzoso, D. Ponce-Balbuena, G. Guerrero-Serna, L. Hou, H. Musa, et al., TGF-beta1, released by myofibroblasts, differentially regulates transcription and function of sodium and potassium channels in adult rat ventricular myocytes, PLoS One, 8 (2013), e55391. |
| [5] | Jan, Trieschmann, Daniel, Bettin, Moritz, Haustein, et al., The Interaction between Adult Cardiac Fibroblasts and Embryonic Stem Cell-Derived Cardiomyocytes Leads to Proarrhythmic Changes in In Vitro Cocultures, Stem cells Int., 2016 (2016), 2936126-2936126. |
| [6] |
J. E. Cartledge, C. Kane, P. Dias, M. Tesfom, L. Clarke, B. Mckee, et al., Functional crosstalk between cardiac fibroblasts and adult cardiomyocytes by soluble mediators, Cardiovasc. Res., 105 (2015), 260-270. doi: 10.1093/cvr/cvu264
|
| [7] |
B. Liau, C. P. Jackman, Y. Li, N. Bursac, Developmental stage-dependent effects of cardiac fibroblasts on function of stem cell-derived engineered cardiac tissues, Sci. Rep., 7 (2017), 42290. doi: 10.1038/s41598-017-18338-2
|
| [8] |
P. M. Boyle, T. V. Karathanos, N. A. Trayanova, Cardiac Optogenetics: 2018, JACC Clin. Electrophysiol., 4 (2018), 155-167. doi: 10.1016/j.jacep.2017.12.006
|
| [9] |
T. A. Quinn, P. Camelliti, E. A. Rog-Zielinska, U. Siedlecka, T. Poggioli, E. T. O'Toole, et al., Electrotonic coupling of excitable and nonexcitable cells in the heart revealed by optogenetics, Proc. Natl. Acad. Sci. U. S. A., 113 (2016), 14852-14857. doi: 10.1073/pnas.1611184114
|
| [10] | J. Yu, P. M. Boyle, A. Klimas, J. C. Williams, N. Trayanova, E. Entcheva, OptoGap: an optogenetics-enabled assay for quantification of cell-cell coupling in multicellular cardiac tissue, bioRxiv, (2017), 171397. |
| [11] |
G. Kostecki, Y. Shi, C. S. Chen, D. H. Reich, L. Tung, Optogenetic currents in myofibroblasts acutely alters electrophysiology and conduction of co-cultured cardiomyocytes, Sci. Rep., 11 (2021), 4430. doi: 10.1038/s41598-021-83398-4
|
| [12] |
J. C. Williams, J. Xu, Z. Lu, A. Klimas, X. Chen, C. M. Ambrosi, et al., Computational optogenetics: empirically-derived voltage- and light-sensitive channelrhodopsin-2 model, PLoS Comput. Biol., 9 (2013), e1003220. doi: 10.1371/journal.pcbi.1003220
|
| [13] |
Z. Jia, V. Valiunas, Z. Lu, H. Bien, H. Liu, H. Z. Wang, et al., Stimulating cardiac muscle by light: cardiac optogenetics by cell delivery, Circ. Arrhythm. Electrophysiol., 4 (2011), 753-760. doi: 10.1161/CIRCEP.111.964247
|
| [14] |
O. J. Abilez, J. Wong, R. Prakash, K. Deisseroth, C. K. Zarins, E. Kuhl, Multiscale computational models for optogenetic control of cardiac function, Biophys. J., 101 (2011), 1326-1334. doi: 10.1016/j.bpj.2011.08.004
|
| [15] |
R. F. Ribeiro, Jr., F. F. Potratz, B. M. Pavan, L. Forechi, F. L. Lima, J. Fiorim, et al., Carvedilol prevents ovariectomy-induced myocardial contractile dysfunction in female rat, PLoS One, 8 (2013), e53226. doi: 10.1371/journal.pone.0053226
|
| [16] |
A. Reed, P. Kohl, R. Peyronnet, Molecular candidates for cardiac stretch-activated ion channels, Glob. Cardiol. Sci. Pract., 2014 (2014), 9-25. doi: 10.5339/gcsp.2014.9
|
| [17] |
N. A. Balakina-Vikulova, A. Panfilov, O. Solovyova, L. B. Katsnelson, Mechano-calcium and mechano-electric feedbacks in the human cardiomyocyte analyzed in a mathematical model, J. Physiol. Sci., 70 (2020), 12. doi: 10.1186/s12576-020-00741-6
|
| [18] |
R. L. Kaufmann, M. J. Lab, R. Hennekes, H. Krause, Feedback interaction of mechanical and electrical events in the isolated mammalian ventricular myocardium (cat papillary muscle), Pflugers Arch., 324 (1971), 100-123. doi: 10.1007/BF00592656
|
| [19] |
M. M. Maleckar, J. L. Greenstein, W. R. Giles, N. A. Trayanova, K+ current changes account for the rate dependence of the action potential in the human atrial myocyte, Am. J. Physiol. Heart Circ. Physiol., 297 (2009), H1398-1410. doi: 10.1152/ajpheart.00411.2009
|
| [20] | S. Coveney, R. H. Clayton, Sensitivity and Uncertainty Analysis of Two Human Atrial Cardiac Cell Models Using Gaussian Process Emulators, Front. Physiol., 11 (2020), 364. |
| [21] |
A. Muszkiewicz, X. Liu, A. Bueno-Orovio, B. A. J. Lawson, K. Burrage, B. Casadei, et al., From ionic to cellular variability in human atrial myocytes: an integrative computational and experimental study, Am. J. Physiol. Heart Circ. Physiol., 314 (2018), H895-H916. doi: 10.1152/ajpheart.00477.2017
|
| [22] |
H. Zhan, J. Zhang, A. Jiao, Q. Wang, Stretch-activated current in human atrial myocytes and Na(+) current and mechano-gated channels' current in myofibroblasts alter myocyte mechanical behavior: a computational study, Biomed. Eng. Online, 18 (2019), 104. doi: 10.1186/s12938-019-0723-5
|
| [23] |
T. Sulman, L. B. Katsnelson, O. Solovyova, V. S. Markhasin, Mathematical modeling of mechanically modulated rhythm disturbances in homogeneous and heterogeneous myocardium with attenuated activity of na+ -k+ pump, Bull. Math. Biol., 70 (2008), 910-949. doi: 10.1007/s11538-007-9285-y
|
| [24] | A. Khokhlova, P. Konovalov, G. Iribe, O. Solovyova, L. Katsnelson, The Effects of Mechanical Preload on Transmural Differences in Mechano-Calcium-Electric Feedback in Single Cardiomyocytes: Experiments and Mathematical Models, Front. Physiol., 11 (2020), 171. |
| [25] |
O. Solovyova, L. Katsnelson, S. Guriev, L. Nikitina, Y. Protsenko, S. Routkevitch, et al., Mechanical inhomogeneity of myocardium studied in parallel and serial cardiac muscle duplexes: experiments and models, Chaos Solitons Fractals, 13 (2002), 1685-1711. doi: 10.1016/S0960-0779(01)00175-8
|
| [26] |
A. Nygren, C. Fiset, L. Firek, J. W. Clark, D. S. Lindblad, R. B. Clark, et al., Mathematical model of an adult human atrial cell: the role of K+ currents in repolarization, Circ. Res., 82 (1998), 63-81. doi: 10.1161/01.RES.82.1.63
|
| [27] |
K. A. MacCannell, H. Bazzazi, L. Chilton, Y. Shibukawa, R. B. Clark, W. R. Giles, A mathematical model of electrotonic interactions between ventricular myocytes and fibroblasts, Biophys. J., 92 (2007), 4121-4132. doi: 10.1529/biophysj.106.101410
|
| [28] |
M. M. Maleckar, J. L. Greenstein, W. R. Giles, N. A. Trayanova, Electrotonic coupling between human atrial myocytes and fibroblasts alters myocyte excitability and repolarization, Biophys. J., 97 (2009), 2179-2190. doi: 10.1016/j.bpj.2009.07.054
|
| [29] |
V. Izakov, L. B. Katsnelson, F. A. Blyakhman, V. S. Markhasin, T. F. Shklyar, Cooperative effects due to calcium binding by troponin and their consequences for contraction and relaxation of cardiac muscle under various conditions of mechanical loading, Circ. Res., 69 (1991), 1171-1184. doi: 10.1161/01.RES.69.5.1171
|
| [30] |
C. F. Vahl, A. Bonz, T. Timek, S. Hagl, Intracellular calcium transient of working human myocardium of seven patients transplanted for congestive heart failure, Circ. Res., 74 (1994), 952-958. doi: 10.1161/01.RES.74.5.952
|
| [31] |
C. F. Vahl, T. Timek, A. Bonz, N. Kochsiek, H. Fuchs, L. Schaffer, et al., Myocardial length-force relationship in end stage dilated cardiomyopathy and normal human myocardium: analysis of intact and skinned left ventricular trabeculae obtained during 11 heart transplantations, Basic Res. Cardiol., 92 (1997), 261-270. doi: 10.1007/BF00788521
|
| [32] |
G. Hasenfuss, L. A. Mulieri, B. J. Leavitt, P. D. Allen, J. R. Haeberle, N. R. Alpert, Alteration of contractile function and excitation-contraction coupling in dilated cardiomyopathy, Circ. Res., 70 (1992), 1225-1232. doi: 10.1161/01.RES.70.6.1225
|
| [33] |
P. M. Boyle, T. V. Karathanos, E. Entcheva, N. A. Trayanova, Computational modeling of cardiac optogenetics: Methodology overview & review of findings from simulations, Comput. Biol. Med., 65 (2015), 200-208. doi: 10.1016/j.compbiomed.2015.04.036
|
| [34] |
U. Nussinovitch, R. Shinnawi, L. Gepstein, Modulation of cardiac tissue electrophysiological properties with light-sensitive proteins, Cardiovasc. Res., 102 (2014), 176-187. doi: 10.1093/cvr/cvu037
|
| [35] |
P. M. Boyle, J. C. Williams, C. M. Ambrosi, E. Entcheva, N. A. Trayanova, A comprehensive multiscale framework for simulating optogenetics in the heart, Nat. Commun., 4 (2013), 2370. doi: 10.1038/ncomms3370
|
| [36] |
C. M. Ambrosi, A. Klimas, J. Yu, E. Entcheva, Cardiac applications of optogenetics, Prog. Biophys. Mol. Biol., 115 (2014), 294-304. doi: 10.1016/j.pbiomolbio.2014.07.001
|
| [37] |
J. C. Williams, E. Entcheva, Optogenetic versus Electrical Stimulation of Human Cardiomyocytes: Modeling Insights, Biophys. J., 108 (2015), 1934-1945. doi: 10.1016/j.bpj.2015.03.032
|
| [38] |
T. V. Karathanos, P. M. Boyle, N. A. Trayanova, Optogenetics-enabled dynamic modulation of action potential duration in atrial tissue: feasibility of a novel therapeutic approach, Europace, 16 (2014), iv69-iv76. doi: 10.1093/europace/euu250
|
| [39] |
T. Bruegmann, D. Malan, M. Hesse, T. Beiert, C. J. Fuegemann, B. K. Fleischmann, et al., Optogenetic control of heart muscle in vitro and in vivo, Nat. Methods, 7 (2010), 897-900. doi: 10.1038/nmeth.1512
|
| [40] |
A. B. Arrenberg, D. Y. Stainier, H. Baier, J. Huisken, Optogenetic control of cardiac function, Science, 330 (2010), 971-974. doi: 10.1126/science.1195929
|
| [41] |
Y. Xie, A. Garfinkel, J. N. Weiss, Z. Qu, Cardiac alternans induced by fibroblast-myocyte coupling: mechanistic insights from computational models, Am. J. Physiol. Heart Circ. Physiol., 297 (2009), H775-784. doi: 10.1152/ajpheart.00341.2009
|
| [42] |
O. Solovyova, N. Vikulova, L. B. Katsnelson, V. S. Markhasin, P. J. Noble, A. Garny, et al., Mechanical interaction of heterogeneous cardiac muscle segments in silico: effects on Ca2+ handling and action potential, Int. J. Bifurcat. Chaos, 13 (2003), 3757-3782. doi: 10.1142/S0218127403008983
|
| [43] |
L. B. Katsnelson, L. V. Nikitina, D. Chemla, O. Solovyova, C. Coirault, Y. Lecarpentier, et al., Influence of viscosity on myocardium mechanical activity: a mathematical model, J. Theor. Biol., 230 (2004), 385-405. doi: 10.1016/j.jtbi.2004.05.007
|
| [44] |
Y. Lecarpentier, A. Waldenstrom, M. Clergue, D. Chemla, P. Oliviero, J. L. Martin, et al., Major alterations in relaxation during cardiac hypertrophy induced by aortic stenosis in guinea pig, Circ. Res., 61 (1987), 107-116. doi: 10.1161/01.RES.61.1.107
|
| [45] |
M. Zabel, B. S. Koller, F. Sachs, M. R. Franz, Stretch-induced voltage changes in the isolated beating heart: importance of the timing of stretch and implications for stretch-activated ion channels, Cardiovasc. Res., 32 (1996), 120-130. doi: 10.1016/S0008-6363(96)00089-2
|
| [46] |
T. Zeng, G. C. Bett, F. Sachs, Stretch-activated whole cell currents in adult rat cardiac myocytes, Am. J. Physiol. Heart Circ. Physiol., 278 (2000), H548-557. doi: 10.1152/ajpheart.2000.278.2.H548
|
| [47] | S. N. Healy, A. D. McCulloch, An ionic model of stretch-activated and stretch-modulated currents in rabbit ventricular myocytes, Europace, 7 (2005), 128-134. |
| [48] | S. A. Park, S. R. Lee, L. Tung, D. T. Yue, Optical mapping of optogenetically shaped cardiac action potentials, Sci. Rep., 4 (2014), 6125. |
 mbe-18-06-414 S1.pdf mbe-18-06-414 S1.pdf |
 |
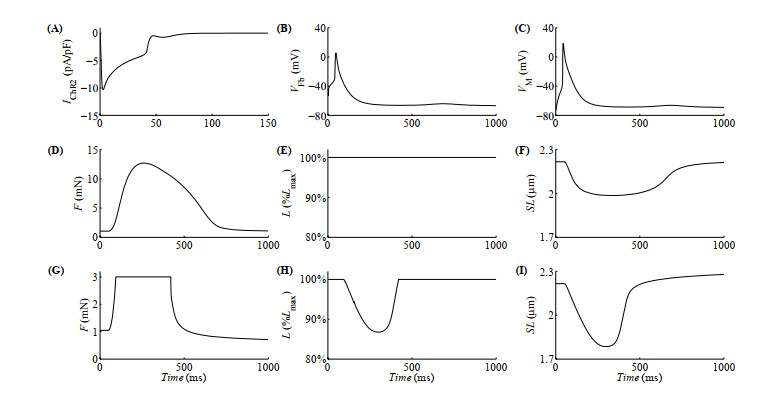
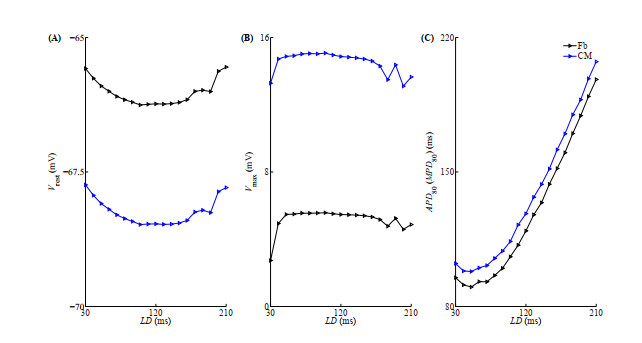
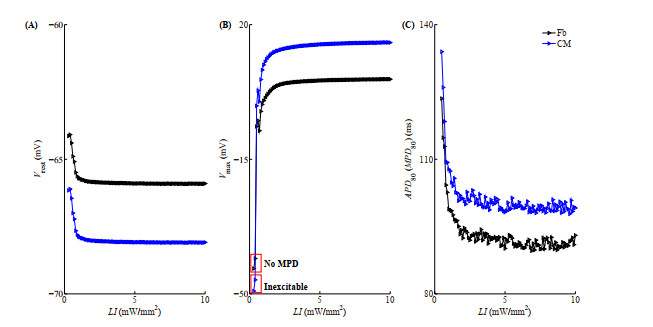
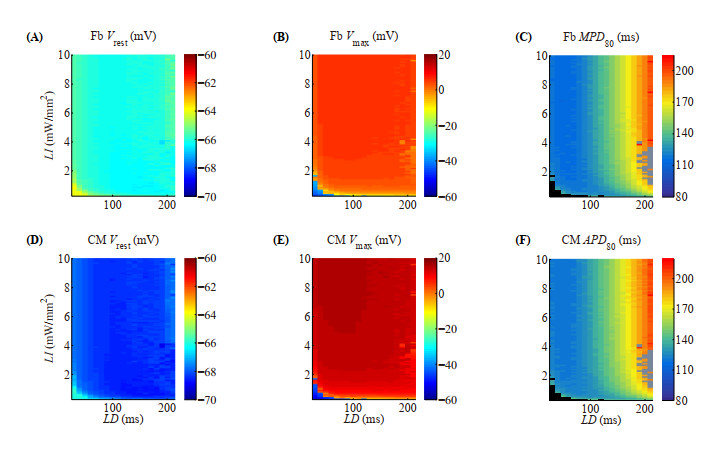
Figures(10)

Heqing Zhan, Zefeng Wang, Jialun Lin, Yuanbo Yu, Ling Xia. Optogenetic actuation in ChR2-transduced fibroblasts alter excitation-contraction coupling and mechano-electric feedback in coupled cardiomyocytes: a computational modeling study[J]. Mathematical Biosciences and Engineering, 2021, 18(6): 8354-8373. doi: 10.3934/mbe.2021414







 DownLoad:
DownLoad: