Citation: Jian Ren, Rui Xu, Liangchen Li. Global stability of an HIV infection model with saturated CTL immune response and intracellular delay[J]. Mathematical Biosciences and Engineering, 2021, 18(1): 57-68. doi: 10.3934/mbe.2021003

| [1] |
A. M. Elaiw, Global properties of a class of HIV models, Nonlinear Anal. RWA, 11 (2010), 2253-2263. doi: 10.1016/j.nonrwa.2009.07.001

|
| [2] |
L. Wang, M. Y. Li, Mathematical analysis of the global dynamics of a model for HIV infection of CD4+ T cells, Math. Biosci., 200 (2006), 44-57. doi: 10.1016/j.mbs.2005.12.026
|
| [3] |
L. Cai, X. Li, M. Ghosh, B. Guo, Stability analysis of an HIV/AIDS epidemic model with treatment, J. Comput. Appl. Math., 229 (2009), 313-323. doi: 10.1016/j.cam.2008.10.067
|
| [4] | J. Xu, Y. Geng, Y. Zhou, Global dynamics for an age-structured HIV virus infection model with cellular infection and antiretroviral therapy, Appl. Math. Comput., 305 (2017), 62-83. |
| [5] |
M. A. Nowak, S. Bonhoeffer, G. M. Shaw, R. M. May, Anti-viral drug treatment: dynamics of resistance in free virus and infected cell populations, J. Theor. Biol., 184 (1997), 203-217. doi: 10.1006/jtbi.1996.0307
|
| [6] |
M. Y. Li, L. Wang, Backward bifurcation in a mathematical model for HIV infection in vivo with anti-retroviral treatment, Nonlinear Anal. RWA, 17 (2014), 147-160. doi: 10.1016/j.nonrwa.2013.11.002
|
| [7] |
G. P. Samanta, Permanence and extinction of a nonautonomous HIV/AIDS epidemic model with distributed time delay, Nonlinear Anal. RWA, 12 (2011), 1163-1177. doi: 10.1016/j.nonrwa.2010.09.010
|
| [8] |
M. A. Nowak, C. R. M. Bangham, Population dynamics of immune responses to persistent viruses, Science, 272 (1996), 74-79. doi: 10.1126/science.272.5258.74
|
| [9] | R. De Boer, Which of our modeling predictions are robust?, PLoS Comput. Biol., 8 (2012), e1002593. |
| [10] | C. Jiang, W. Wang, Complete classification of global dynamics of a virus model with immune responses, Discrete Contin. Dyn. Syst. Ser. B, 19 (2014), 1087-1103. |
| [11] |
A. V. M. Herz, S. Bonhoeffer, R. M. Anderson, R. M. May, M. A. Nowak, Viral dynamics in vivo: Limitations on estimates of intracellular delay and virus decay, Proc. Natl. Acad. Sci. USA, 93 (1996), 7247-7251. doi: 10.1073/pnas.93.14.7247
|
| [12] |
A. M. Elaiw, I. A. Hassanien, S. A. Azoz, Global stability of HIV infection models with intracellular delays, J. Korean Math. Soc., 49 (2012), 779-794. doi: 10.4134/JKMS.2012.49.4.779
|
| [13] |
B. Reddy, J. Yin, Quantitative intracellular kinetics of HIV type 1, AIDS Res. Hum. Retrovir., 15 (1999), 273-283. doi: 10.1089/088922299311457
|
| [14] | A. S. Perelson, A. U. Neumann, M. Markowitz, J. M. Leonard, D. D. Ho, HIV-1 dynamics in vivo: Virion clearance rate, infected cell life-span, and viral generation time, Science, 271 (1996), 1582-1586. |
| [15] |
J. E. Mittler, B. Sulzer, A. U. Neumann, A. S. Perelson, Influence of delayed viral production on viral dynamics in HIV-1 infected patients, Math. Biosci., 152 (1998), 143-163. doi: 10.1016/S0025-5564(98)10027-5
|
| [16] |
D. Ebert, C. D. Zschokke-Rohringer, H. J. Carius, Dose effects and density-dependent regulation of two microparasites of Daphnia magna, Oecologia, 122 (2000), 200-209. doi: 10.1007/PL00008847
|
| [17] |
R. R. Regoes, D. Ebert, S. Bonhoeffer, Dose-dependent infection rates of parasites produce the Allee effect in epidemiology, Proc. R. Soc. Lond. Ser. B, 269 (2002), 271-279. doi: 10.1098/rspb.2001.1816
|
| [18] |
X. Song, A. U. Neumann, Global stability and periodic solution of the viral dynamics, J. Math. Anal. Appl., 329 (2007), 281-297. doi: 10.1016/j.jmaa.2006.06.064
|
| [19] | J. K. Hale, S. Verduyn Lunel, Introduction to Functional Differential Equations, Springer, New York, 1993. |
| [20] |
P. van den Driessche, J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002), 29-48. doi: 10.1016/S0025-5564(02)00108-6
|
| [21] |
M. Stafford, L. Corey, Y. Cao, E. Daar, D. Ho, A. Perelson, Modeling plasma virus concentration during primary HIV infection, J. Theor. Biol., 203 (2000), 285-301. doi: 10.1006/jtbi.2000.1076
|
| [22] |
K. Pawelek, S. Liu, F. Pahlevani, L. Rong, A model of HIV-1 infection with two time delays: Mathematical analysis and comparison with patient data, Math. Biosci., 235 (2012), 98-109. doi: 10.1016/j.mbs.2011.11.002
|
| [23] | J. Wang, M. Guo, X. Liu, Z. Zhao, Threshold dynamics of HIV-1 virus model with cell-to-cell transmission, cell-mediated immune responses and distributed delay, Appl. Math. Comput., 291 (2016), 149-161. |
| [24] |
S. Marino, I. B. Hogue, C. J. Ray, A methodology for performing global uncertainty and sensitivity analysis in systems biology, J. Theor. Biol., 254 (2008), 178-196. doi: 10.1016/j.jtbi.2008.04.011
|
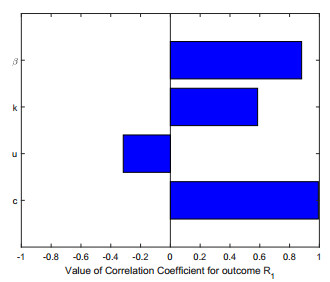
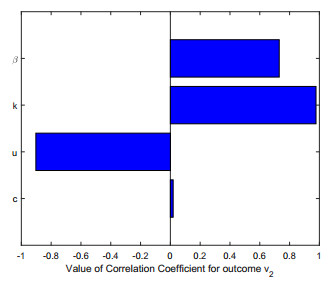
Figures(2)

Jian Ren, Rui Xu, Liangchen Li. Global stability of an HIV infection model with saturated CTL immune response and intracellular delay[J]. Mathematical Biosciences and Engineering, 2021, 18(1): 57-68. doi: 10.3934/mbe.2021003







 DownLoad:
DownLoad: