Citation: Xiaohong Tian, Rui Xu, Jiazhe Lin. Mathematical analysis of an age-structured HIV-1 infection model with CTL immune response[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 7850-7882. doi: 10.3934/mbe.2019395

| [1] | H. D. Kwon, Optimal treatment strategies derived from a HIV model with drug-resistant mutants, Appl. Math. Comput., 188 (2007), 1193–1204. |
| [2] | S. Bonhoeffer, R. M. May, G. M. Shaw, et al., Virus dynamics and drug therapy, Proc. Natl. Acad. Sci. USA, 94 (1997), 6971–6976. |
| [3] | D. D. Ho, A. U. Neumann, A. S. Perelson, et al., Rapid turnover of plasma virions and CD4+ lymphocytes in HIV-1 infection, Nature, 373 (1995), 123–126. |
| [4] | M. A. Nowak, R. M. Anderson, M. C. Boerlijst, et al., HIV-1 evolution and disease progression, Science, 274 (1996), 1008–1011. |
| [5] | M. A. Nowak, S. Bonhoeffer, G. M. Shaw, et al., Anti-viral drug treatment: Dynamics of resistance in free virus and infected cell populations, J. Theor. Biol., 184 (1997), 203–217. |
| [6] | A. S. Perelson and P. W. Nelson, Mathematical analysis of HIV-1 dynamics in vivo, SIAM Rev., 41 (1999), 3–44. |
| [7] | A. S. Perelson, D. E. Kirschner and R. De Boer, Dynamics of HIV infection of CD4+T cells, Math. Biosci., 114 (1993), 81–125. |
| [8] | A. S. Perelson, A. U. Neumann, M. Markowitz, et al., HIV-1 dynamics in vivo: Virion clearance rate, infected cell life-span, and viral generation time, Science, 271 (1996), 1582–1586. |
| [9] | P. W. Nelson, M. A. Gilchrist, D. Coombs, et al., An age-structured model of HIV infection that allows for variations in the production rate of viral particles and the death rate of productively infected cells, Math. Biosci. Eng., 1 (2004), 267–288. |
| [10] | G. Huang, X. Liu and Y. Takeuchi, Lyapunov functions and global stability for age-structured HIV infection model, SIAM J. Appl. Math., 72 (2012), 25–38. |
| [11] | L. Rong, Z. Feng and A. S. Perelson, Mathematical analysis of age-structured HIV-1 dynamics with combination antiretroviral therapy, SIAM J. Appl. Math., 67 (2007), 731–756. |
| [12] | R. Xu, X. Tian and S. Zhang, An age-structured within-host HIV-1 infection model with virus-to-cell and cell-to-cell transmissions, J. Biol. Dyn., 12 (2017), 89–117. |
| [13] | G. W. Suryawanshi and A. Hoffmann, A multi-scale mathematical modeling framework to investigate anti-viral therapeutic opportunities in targeting HIV-1 accessory proteins, J. Theor. Biol., 386 (2015), 89–104. |
| [14] | J. Xu, Y. Geng and Y. Zhou, Global dynamics for an age-structured HIV virus infection model with cellular infection and antiretroviral therapy, Appl. Math. Comput., 305 (2017), 62–83. |
| [15] | J. Wang, J. Lang and X. Zou, Analysis of an age structured HIV infection model with virus-to-cell infection and cell-to-cell transmission, Nonlinear Anal. RWA, 34 (2017), 75–96. |
| [16] | J. Wang, R. Zhang and T. Kuniya, Global dynamics for a class of age-infection HIV models with nonlinear infection rate, J. Math. Anal. Appl., 432 (2015), 289–313. |
| [17] | A. Alshorman, C. Samarasinghe, W. Lu, et al., An HIV model with age-structured latently infected cells, J. Biol. Dyn., 11 (2017), 192–215. |
| [18] | Z. Liu and Z. Li, Molecular imaging in tracking tumor-specific cytotoxic T lymphocytes (CTLs), Theranostics, 4 (2014), 990–1001. |
| [19] | P. K. Roy and A. N. Chatterjee, T-cell proliferation in a mathematical model of CTL activity through HIV-1 infection, Proc. World Congr. Eng., 1 (2010), 1–6. |
| [20] | J. Cao, J. McNevin, S. Holte, et al., Comprehensive analysis of human immunodeficiency virus type 1 (HIV-1)-specific gamma interferon-secreting CD8 + T cells in primary HIV-1 infection, J. Virol., 77 (2003), 6867–6878. |
| [21] | C. Browne, Immune response in virus model structured by cell infection-age, Math. Biosci. Eng., 13 (2016), 887–909. |
| [22] | J. Pang, J. Chen, Z. Liu, et al., Local and global stabilities of a viral dynamics model with infection-age and immune response, J. Dyn. Differ. Equ., 31 (2019), 793–813. |
| [23] | R. R. Regoes, D. Ebert and S. Bonhoeffer, Dose-dependent infection rates of parasites produce the Allee effect in epidemiology, Proc. R. Soc. London B, 269 (2002), 271–279. |
| [24] | M. Iannelli, Mathematical Theory of Age-Structured Population Dynamics, Applied Mathematics Monographs, 7, Consiglio Nazionale delle Ricerche (C.N.R), Giardini Pisa, 1995, comitato nazionale per le scienze matematiche. |
| [25] | G. Webb, Theory of Nonlinear Age-Dependent Population Dynamics, Marcel Dekker, New York, 1985. |
| [26] | H. L. Smith and H. R. Thieme, Dynamical Systems and Population Persistence, American Mathematical Society, Providence, 2011. |
| [27] | J. Yang, R. Xu and X. Luo, Dynamical analysis of an age-structured multi-group SIVS epidemic model, Math. Biosci. Eng., 16 (2019), 636–666. |
| [28] | D. Burg, L. Rong, A. U. Neumann, et al., Mathematical modeling of viral kinetics under immune control during primary HIV-1infection, J. Theor. Biol., 259 (2009), 751–759. |
| [29] | N. M. Dixit and A. S. Perelson, Complex patterns of viral load decay under antiretroviral therapy: Influence of pharmacokinetics and intracellular delay, J. Theor. Biol., 226 (2004), 95–109. |
| [30] | M. Markowitz, M. Louie, A. Hurley, et al., A novel antiviral intervention results in more accurate assessment of human immunodeficiency virus type 1 replication dynamics and t-cell decay in vivo, J. Virol., 77 (2003), 5037–5038. |
| [31] | B. Ramratnam, S. Bonhoeffer, J. Binley, et al., Rapid production and clearance of HIV-1 and hepatitis C virus assessed by large volume plasma apheresis, Lancet, 354 (1999), 1782–1785. |
| [32] | J. Wang, M. Guo, X Liu, et al., Threshold dynamics of HIV-1 virus model with cell-to-cell transmission, cell-mediated immune responses and distributed delay, Appl. Math. Comput., 291 (2016), 149–161. |
| [33] | X. Zhou and J. Cui, Global stability of the viral dynamics with Crowley-Martin functional response, Bull. Korean Math. Soc., 48 (2011), 555–574. |
| [34] | M. Iannelli and F. Milner, The Basic Approach to Age-Structured Population Dynamics: Models, Methods and Numerics, Springer, 2017. |
| [35] | J. K. Hale and P. Waltman, Persistence in infinite-dimensional systems, SIAM J. Math. Anal., 20 (1989), 388–395. |
| [36] | P. Magal, Compact attractors for time periodic age-structured population models, Electron. J. Differ. Equ., 65 (2001), 1–35. |
| [37] | P. Magal, C. C. McCluskey and G. F. Webb, Lyapunov functional and global asymptotic stability for an infection-age model, Appl. Anal., 89 (2010), 1109–1140. |
| [38] | R. Xu, X. Tian and F. Zhang, Global dynamics of a tuberculosis transmission model with age of infection and incomplete treatment, Adv. Differ. Equ., 242 (2017), 1–34. |
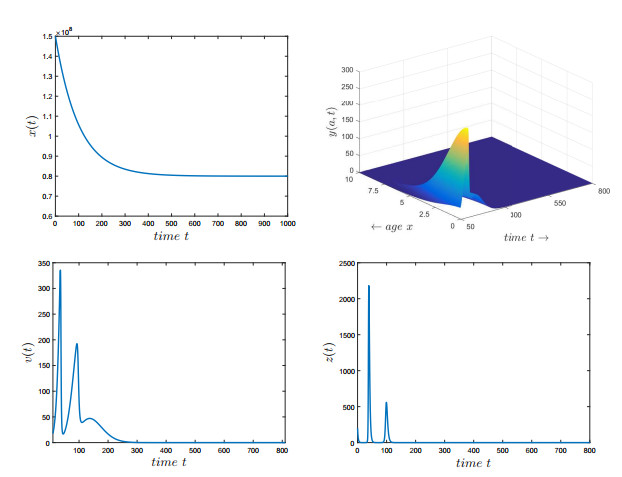
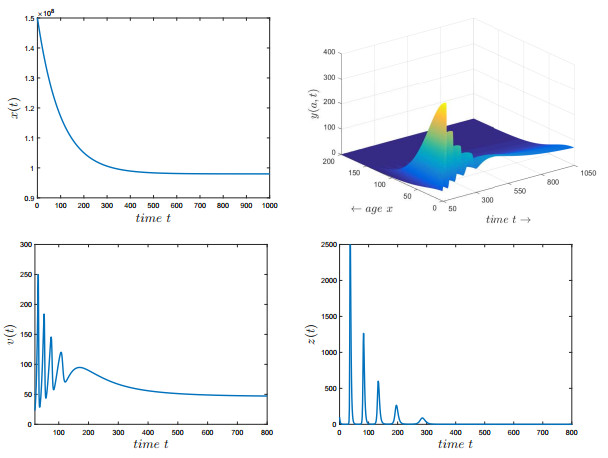
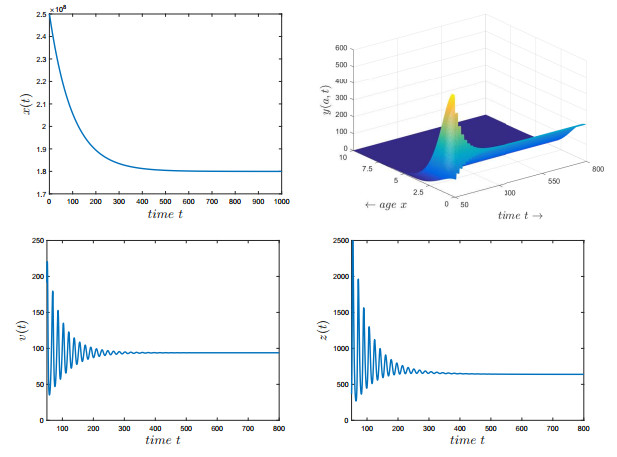
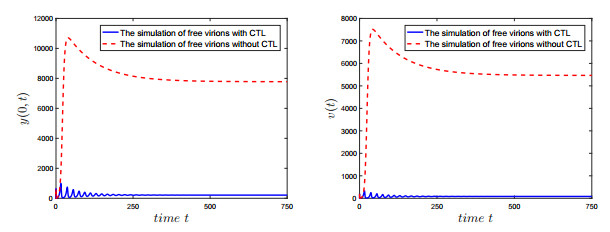
Figures(7) / Tables(2)

Xiaohong Tian, Rui Xu, Jiazhe Lin. Mathematical analysis of an age-structured HIV-1 infection model with CTL immune response[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 7850-7882. doi: 10.3934/mbe.2019395







 DownLoad:
DownLoad: