Citation: Yan Wang, Tingting Zhao, Jun Liu. Viral dynamics of an HIV stochastic model with cell-to-cell infection, CTL immune response and distributed delays[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 7126-7154. doi: 10.3934/mbe.2019358

| [1] | M. A. Nowak and C. Bangham, Population dynamics of immune response to persistent viruses, Science, 272 (1996), 74–79. |
| [2] | H. L. Smith and P. D. Leenheer, Virus dynamics: a global analysis, SIAM J. Appl. Math., 63 (2003), 1313–1327. |
| [3] | G. Huang, Y. Takeuchi and W. Ma, Lyapunov functionals for delay differential equations model of viral infections, SIAM J. Appl. Math., 70 (2010), 2693–2708. |
| [4] | M. Y. Li and H. Shu, Impact of intracellular delays and target-cell dynamics on in vivo viral infections, SIAM J. Appl. Math., 70 (2010), 2434–2448. |
| [5] | M. A. Nowak and R. M. May, Virus dynamics: mathematical principles of immunology and virology, Oxford University, Oxford, 2000. |
| [6] | A. S. Perelson and P. W. Nelson, Mathematical analysis of HIV-1 dynamics in vivo, SIAM Rev., 41 (1999), 3–44. |
| [7] | Y. Wang, Y. Zhou, F. Brauer, et al., Viral dynamics model with CTL immune response incorporating antiretroviral therapy, J. Math. Biol., 67 (2013), 901–934. |
| [8] | Y. Wang, Y. Zhou, J. Wu, et al., Oscillatory viral dynamics in a delayed HIV pathogenesis model, Math. Biosci., 219 (2009), 104–112. |
| [9] | P. Zhong, L. M. Agosto, J. B. Munro, et al., Cell-to-cell transmission of viruses, Curr. Opin. Virol., 3 (2013), 44–50. |
| [10] | S. Gummuluru, C. M. Kinsey and M. Emerman, An in vitro rapid-turnover assay for human immunodeficiency virus type 1 replication selects for cell-to-cell spread of virus, J. Virol., 74 (2000), 10882–10891. |
| [11] | H. Sato, J. Orenstein, D. Dimitrov, et al., Cell-to-cell spread of HIV-1 occurs within minutes and may not involve the participation of virus particles, Virology, 186 (1992), 712–724. |
| [12] | A. Sigal, J. T. Kim, A. B. Balazs, et al., Cell-to-cell spread of HIV permits ongoing replication despite antiretroviral therapy, Nature, 477 (2011), 95–98. |
| [13] | R. V. Culshaw, S. Ruan and G. Webb, A mathematical model of cell-to-cell spread of HIV-1 that includes a time delay, J. Math. Biol., 46 (2003), 425–444. |
| [14] | X. Lai and X. Zou, Modeling HIV-1 virus dynamics with both virus-to-cell infection and cell-to-cell transmission, SIAM J. Appl. Math., 74 (2014), 898–917. |
| [15] | X. Lai and X. Zou, Modeling cell-to-cell spread of HIV-1 with logistic target cell growth, J. Math. Anal. Appl., 426 (2015), 563–584. |
| [16] | F. Li and J. Wang, Analysis of an HIV infection model with logistic target-cell growth and cell-to-cell transmission, Chaos. Soliton. Fract., 81 (2015), 136–145. |
| [17] | X. Wang, S. Tang, X. Song, et al., Mathematical analysis of an HIV latent infection model including both virus-to-cell infection and cell-to-cell transmission, J. Biol. Dynam., 11 (2017), 455–483. |
| [18] | S. S. Chen, C. Y. Cheng and Y. Takeuchi, Stability analysis in delayed within-host viral dynamics with both viral and cellular infections, J. Math. Anal. Appl., 442 (2016), 642–672. |
| [19] | Y. Nakata, Global dynamics of a cell mediated immunity in viral infection models with distributed delays. J. Math. Anal. Appl., 375 (2011), 14–27. |
| [20] | T. Nicoleta, Drug therapy model with time delays for HIV infection with virus-to-cell and cell-to- cell transmissions, J. Appl. Math. Comput., 59 (2019), 677–691. |
| [21] | J. Xu and Y. Zhou, Bifurcation analysis of HIV-1 infection model with cell-to-cell transmission and immune response delay, Math. Biosci. Eng., 13 (2017), 343–367. |
| [22] | H. Shu, Y. Chen and L. Wang, Impacts of the cell-free and cell-to-cell infection modes on viral dynamics, J. Dyn. Differ. Equ., 30 (2018), 1817–1836. |
| [23] | Y. Yang, L. Zou and S. Ruan, Global dynamics of a delayed within-host viral infection model with both virus-to-cell and cell-to-cell transmissions, Math. Biosci., 270 (2015), 183–191. |
| [24] | J. Wang, M. Guo, X. Liu, et al., Threshold dynamics of HIV-1 virus model with cell-to-cell transmission, cell-mediated immune responses and distributed delay, Appl. Math. Comput., 291 (2016), 149–161. |
| [25] | Y. Cai, Y. Kang and W. Wang, A stochastic SIRS epidemic model with nonlinear incidence rate, Appl. Math. Comput., 305 (2017), 221–240. |
| [26] | D. Li, J. Cui, M. Liu, et al., The evolutionary dynamics of stochastic epidemic model with nonlinear incidence rate, Bull. Math. Biol., 77 (2015), 1705–1743. |
| [27] | X. Meng, S. Zhao, T. Feng, et al., Dynamics of a novel nonlinear stochastic SIS epidemic model with double epidemic hypothesis, J. Math. Anal. Appl., 433 (2016), 227–242. |
| [28] | Z. Teng and L. Wang, Persistence and extinction for a class of stochastic SIS epidemic models with nonlinear incidence rate, Physica A., 451 (2016), 507–518. |
| [29] | Y. Tan, L. Ning, S. Tang, et al., Optimal threshold density in a stochastic resource management model with pulse intervention, Nat. Resour. Model., (2019), e12220. |
| [30] | H. H. Mcadams and A. Arkin, Stochastic mechanisms in gene expression, Proc. Natl. Acad. Sci. USA, 94 (1997), 814–819. |
| [31] | K. Millerjensen, R. Skupsky, P. S. Shah, et al., Genetic selection for context-dependent stochastic phenotypes: Sp1 and TATA mutations increase phenotypic noise in HIV-1 gene expression, PLos. Comp. Biol., 9 (2013), e1003135. |
| [32] | X. Mao, G. Marion and E. Renshaw, Environmental Brownian noise suppresses explosions in population dynamics, Stoch. Proc. Appl., 97 (2002), 95–110. |
| [33] | N. Dalal, D. Greenhalgh and X. Mao, A stochastic model for internal HIV dynamics, J. Math. Anal. Appl., 341 (2008), 1084–1101. |
| [34] | Z. Huang, Q. Yang and J. Cao, Complex dynamics in a stochastic internal HIV model, Chaos. Soliton. Fract., 44 (2011), 954–963. |
| [35] | H. C. Tuckwell and E. Lecorfec, A stochastic model for early HIV-1 population dynamics, J. Theor. Biol., 195 (1998), 451–463. |
| [36] | Y. Wang, D. Jiang, T. Hayat, et al., A stochastic HIV infection model with T-cell proliferation and CTL immune response, Appl. Math. Comput., 315 (2017), 477–493. |
| [37] | C. Ji, Q. Liu and D. Jiang, Dynamics of a stochastic cell-to-cell HIV-1 model with distributed delay, Physica A., 492 (2018), 1053–1065. |
| [38] | T. Feng, Z. Qiu, X. Meng, et al., Analysis of a stochastic HIV-1 infection model with degenerate diffusion, Appl. Math. Comput., 348 (2019), 437–455. |
| [39] | Q. Liu, D. Jiang, N. Shi, et al., Stationarity and periodicity of positive solutions to stochastic SEIR epidemic models with distributed delay, Discrete Cont. Dyn-B, 22 (2017), 2479–2500. |
| [40] | W. Zuo, D. Jiang, X. Sun, et al., Long-time behaviors of a stochastic cooperative Lotka CVolterra system with distributed delay, Physica A. 506 (2018), 542–559. |
| [41] | X. Ji, S. Yuan, T. Zhang, et al., Stochastic modeling of algal bloom dynamics with delayed nutrient recycling, Math. Biosci. Eng., 16 (2018), 1–24. |
| [42] | N. Macdonald, Time lags in biological models, Lecture Notes in Biomathematics, Springer-Verlag, Heidelberg, 1978. |
| [43] | J. Mittler, B. Sulzer, A. Neumann, et al., Influence of delayed virus production on viral dynamics in HIV-1 infected patients, Math. Biosci., 152 (1998), 143–163. |
| [44] | X. Mao, Stochastic differential equations and applications, 2nd edition, Horwood, Chichester, UK, 2008. |
| [45] | R. Khasminskii, Stochastic stability of differential equations, Sijthoff & Noordhoff, Alphen aan den Rijn, The Netherlands, 1980. |
| [46] | N. Ikeda and S. Watanabe, A comparison theorem for solutions of stochastic differential equations and its applications, Osaka. J. Math., 14 (1977), 619–633. |
| [47] | D. J. Higham, An algorithmic introduction to numerical simulation of stochastic differential equations, SIAM. Rev., 43 (2001), 525–546. |
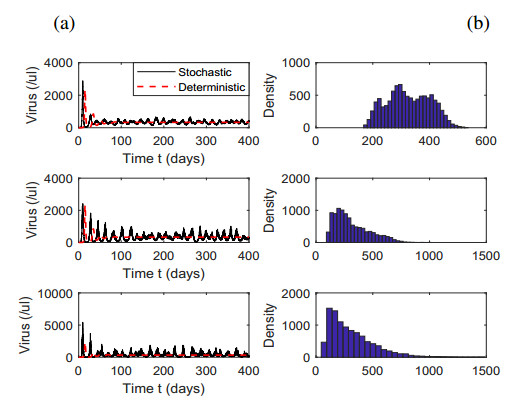
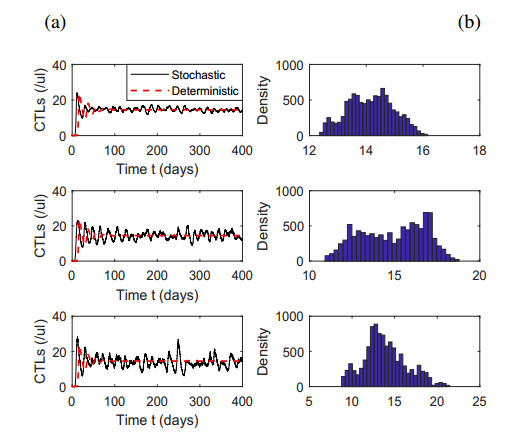
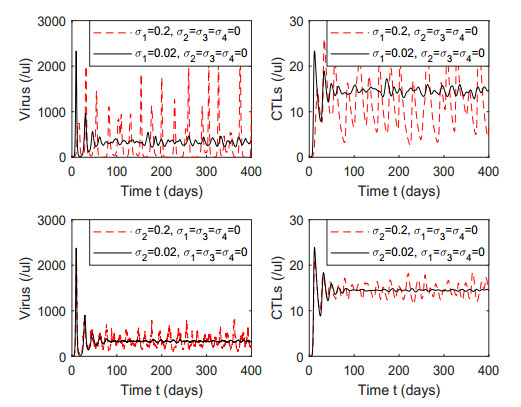
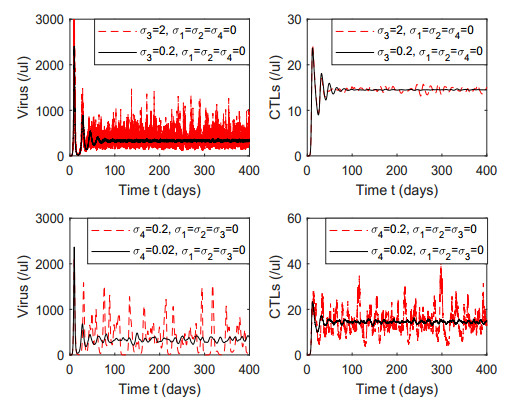
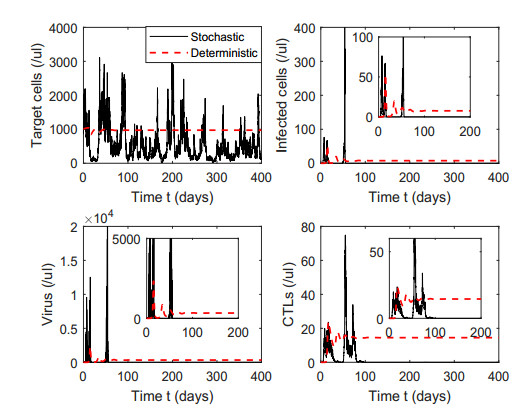
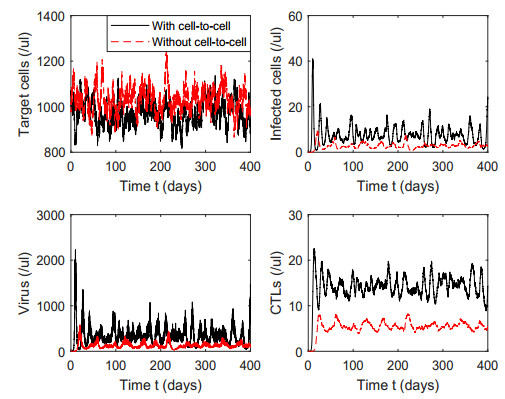
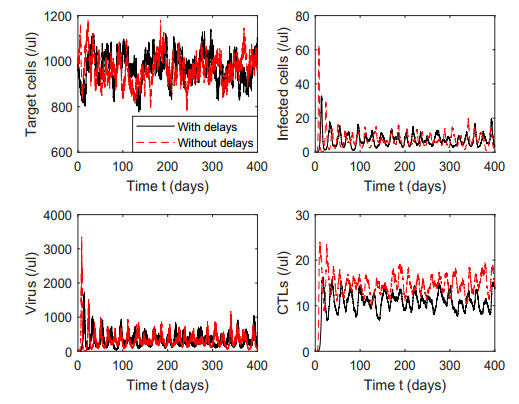
Figures(7) / Tables(1)

Yan Wang, Tingting Zhao, Jun Liu. Viral dynamics of an HIV stochastic model with cell-to-cell infection, CTL immune response and distributed delays[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 7126-7154. doi: 10.3934/mbe.2019358







 DownLoad:
DownLoad: