Citation: Yan Wang, Minmin Lu, Daqing Jiang. Viral dynamics of a latent HIV infection model with Beddington-DeAngelis incidence function, B-cell immune response and multiple delays[J]. Mathematical Biosciences and Engineering, 2021, 18(1): 274-299. doi: 10.3934/mbe.2021014

| [1] |
T. W. Chun, L. Stuyver, S. B. Mizell, L. A. Ehler, J. A. M. Mican, M. Baseler, et al., Presence of an inducible HIV-1 latent reservoir during highly active antiretroviral therapy, Proc. Natl. Acad. Sci., 94 (1997), 13193-13197. doi: 10.1073/pnas.94.24.13193

|
| [2] |
D. Finzi, J. Blankson, J. D. Siliciano, J. B. Margolick, K. Chadwick, T. Pierson, et al., Latent infection of CD4+ T cells provides a mechanism for lifelong persistence of HIV-1, even in patients on effective combination therapy, Nat. Med., 5 (1999), 512-517. doi: 10.1038/8394
|
| [3] |
A. S. Perelson, P. W. Nelson, Mathematical analysis of HIV-1 dynamics in vivo, SIAM Rev., 41 (1999), 3-44. doi: 10.1137/S0036144598335107
|
| [4] | M. A. Nowak, R. M. May, Virus dynamics: mathematical principles of immunology and virology, Oxford University, Oxford, 2000. |
| [5] |
L. Rong, A. S. Perelson, Modeling HIV persistence, the latent reservoir, and viral blips, J. Theor. Biol., 260 (2009), 308-331. doi: 10.1016/j.jtbi.2009.06.011
|
| [6] | L. Rong, A. S. Perelson, Modeling latently infected cell activation: viral and latent reservoir persistence, and viral blips in HIV-infected patients on potent therapy, PLoS Comput. Biol., 5 (2009), 1-18. |
| [7] |
J. A. Deans, S. Cohen, Immunology of malaria, Annu. Rev. Microbiol., 37 (1983), 25-49. doi: 10.1146/annurev.mi.37.100183.000325
|
| [8] |
X. Song, A. Neumann, Global stability and periodic solution of the viral dynamics, J. Math. Anal. Appl., 329 (2007), 281-297. doi: 10.1016/j.jmaa.2006.06.064
|
| [9] |
D. Li, W. Ma, Asymptotic properties of an HIV-1 infection model with time delay, J. Math. Anal. Appl., 335 (2007), 683-691. doi: 10.1016/j.jmaa.2007.02.006
|
| [10] |
G. Huang, W. Ma, Y. Takeuchi, Global properties for virus dynamics model with BeddingtonDeAngelis functional response, Appl. Math. Lett., 22 (2009), 1690-1693. doi: 10.1016/j.aml.2009.06.004
|
| [11] |
G. Huang, W. Ma, Y. Takeuchi, Global analysis for delay virus dynamics model with BeddingtonDeAngelis functional response, Appl. Math. Lett., 24 (2011), 1199-1203. doi: 10.1016/j.aml.2011.02.007
|
| [12] |
H. Miao, Z. Teng, X. Abdurahman, Stability and Hopf bifurcation for a five-dimensional virus infection model with Beddington-DeAngelis incidence and three delays, J. Biol. Dynam., 12 (2018), 146-170. doi: 10.1080/17513758.2017.1408861
|
| [13] |
H. Shu, L. Wang, J. Watmough, Global stability of a nonlinear viral infection model with infinitely distributed intracellular delays and CTL immune responses, SIAM J. Appl. Math., 73 (2013), 1280-1302. doi: 10.1137/120896463
|
| [14] |
J. R. Beddington, Mutual interference between parasites or predators and its effect on searching efficiency, J. Anim. Ecol., 44 (1975), 331-340. doi: 10.2307/3866
|
| [15] |
D. L. DeAngelis, R. A. Goldstein, R. V. O'Neill, A model for trophic interaction, Ecology, 56 (1975), 881-892. doi: 10.2307/1936298
|
| [16] |
A. V. M. Herz, S. Bonhoeffer, R. M. Anderson, R. M. May, M. A. Nowak, Viral dynamics in vivo: limitations on estimates of intracellular delay and virus decay, Proc. Natl. Acad. Sci. USA, 93 (1996), 7247-7251. doi: 10.1073/pnas.93.14.7247
|
| [17] |
H. Liu, J. Zhang, Dynamics of two time delays differential equation model to HIV latent infection, Physica A., 514 (2019), 384-395. doi: 10.1016/j.physa.2018.09.087
|
| [18] | Y. Yang, Y. Dong, Y. Takeuchi, Global dynamics of a latent HIV infection model with general incidence function and multiple delays, Discrete Cont. Dyn. B, 24 (2019), 783-800. |
| [19] |
Y. Wang, M. Lu, J. Liu. Global stability of a delayed virus model with latent infection and Beddington-DeAngelis infection function, Appl. Math. Lett., 107 (2020), 106463. doi: 10.1016/j.aml.2020.106463
|
| [20] |
Y. Wang, F. Brauer, J. Wu, J. M. Heffernan, A delay-dependent model with HIV drug resistance during therapy, J. Math. Anal. Appl., 414 (2014), 514-531. doi: 10.1016/j.jmaa.2013.12.064
|
| [21] |
X. Wang, S. Tang, X. Song, L. Rong, Mathematical analysis of an HIV latent infection model including both virus-to-cell infection and cell-to-cell transmission, J. Biol. Dynam., 11 (2017), 455-483. doi: 10.1080/17513758.2016.1242784
|
| [22] |
Y. Wang, Y. Zhou, J. Wu, J. Heffernan, Oscillatory viral dynamics in a delayed HIV pathogenesis model, Math. Biosci., 219 (2009), 104-112. doi: 10.1016/j.mbs.2009.03.003
|
| [23] |
A. Alshorman, X. Wang, M. J. Meyer, L. Rong, Analysis of HIV models with two time delays, J. Biol. Dynam., 11 (2017), 40-64. doi: 10.1080/17513758.2016.1148202
|
| [24] |
J. Lin, R. Xu, X. Tian, Threshold dynamics of an HIV-1 model with both viral and cellular infections, cell-mediated and humoral immune responses, Math. Biosci. Eng., 16 (2019), 292-319. doi: 10.3934/mbe.2019015
|
| [25] | J. Lin, R. Xu, X. Tian, Threshold dynamics of an HIV-1 virus model with both virus-to-cell and cell-to-cell transmissions, intracellular delay, and humoral immunity, Appl. Math. Comput., 315 (2017), 516-530. |
| [26] |
H. Xiang, L. Feng, H. Huo, Stability of the virus dynamics model with Beddington-DeAngelis functional response and delays, Appl. Math. Model., 37 (2013), 5414-5423. doi: 10.1016/j.apm.2012.10.033
|
| [27] |
X. Song, X. Zhou, X. Zhao, Properties of stability and Hopf bifurcation for a HIV infection model with time delay, Appl. Math. Model., 34 (2010), 1511-1523. doi: 10.1016/j.apm.2009.09.006
|
| [28] |
B. Li, Y. Chen, X. Lu, S. Liu, A delayed HIV-1 model with virus waning term, Math. Biosci. Eng., 13 (2016), 135-157. doi: 10.3934/mbe.2016.13.135
|
| [29] |
J. Xu, Y. Geng, S. Zhang, Global stability and Hopf bifurcation in a delayed viral infection model with cell-to-cell transmission and humoral immune response, Int. J. Bifurcat. Chaos, 29 (2019) 1950161. doi: 10.1142/S021812741950161X
|
| [30] | J. Hale, S. M. Verduyn Lunel, Introduction to functional differential equations, Springer-Verlag, New York, 1993. |
| [31] |
P. van den Driessche, J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002), 29-48. doi: 10.1016/S0025-5564(02)00108-6
|
| [32] |
J. M. Heffernan, R. J. Smith, L. M. Wahl, Perspectives on the basic reproductive ratio, J. R. Soc. Interf., 2 (2005), 281-293. doi: 10.1098/rsif.2005.0042
|
| [33] |
C. C. McCluskey, Complete global stability for an SIR epidemic model with delay-distributed or discrete, Nonlinear Anal. Real, 11 (2010), 55-59. doi: 10.1016/j.nonrwa.2008.10.014
|
| [34] | J. P. LaSalle, The stability of dynamical systems, Regional Conference Series in Applied Mathematics, SIAM Philadelphia, 1976. |
| [35] |
J. M. Heffernan, L. M. Wahl, Natural variation in HIV infection: monte carlo estimates that include CD8 effector cells. J. Theor. Biol. 243 (2006), 191-204. doi: 10.1016/j.jtbi.2006.05.032
|
| [36] |
Y. Wang, Y. Zhou, F. Brauer, J. M. Heffernan, Viral dynamics model with CTL immune response incorporating antiretroviral therapy, J. Math. Biol., 67 (2013), 901-934. doi: 10.1007/s00285-012-0580-3
|
| [37] |
M. A. Nowak, C. Bangham, Population dynamics of immune response to persistent viruses, Science, 272 (1996), 74-79. doi: 10.1126/science.272.5258.74
|
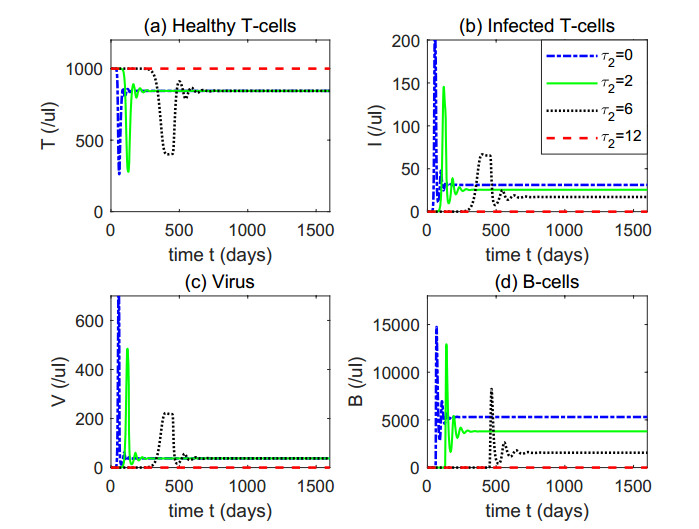
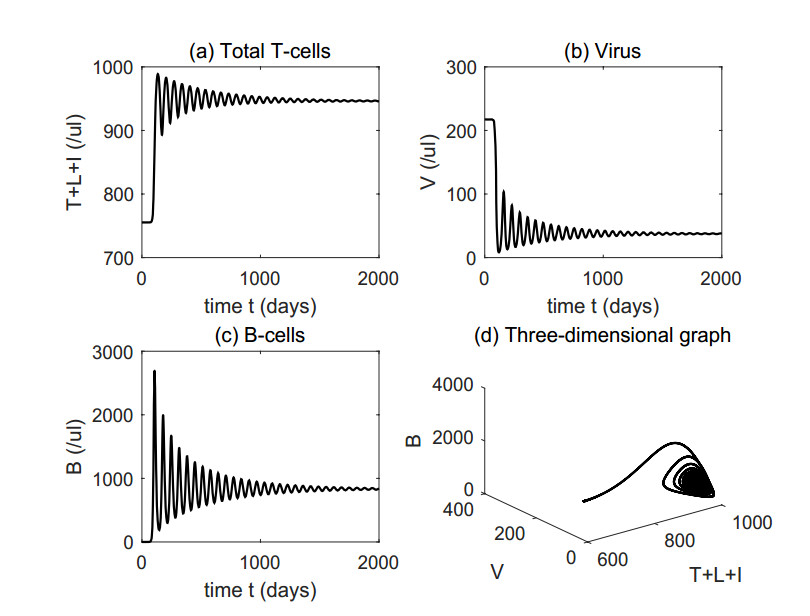
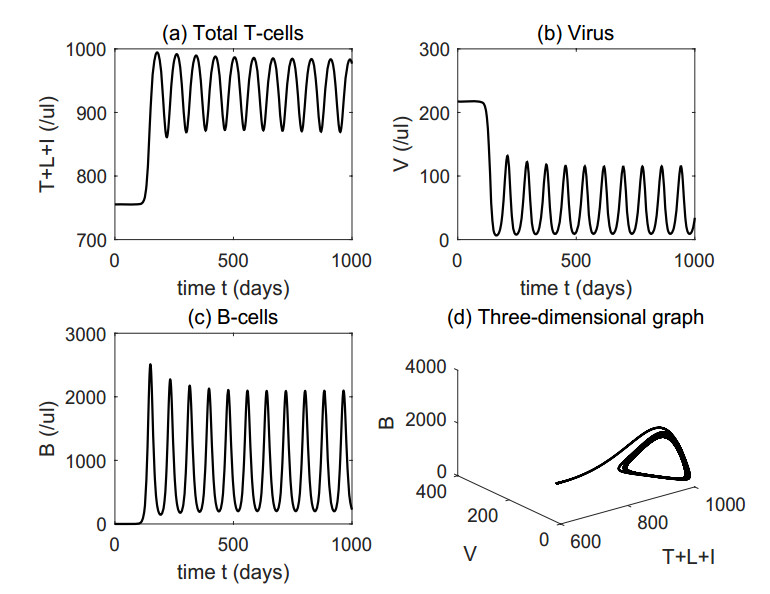
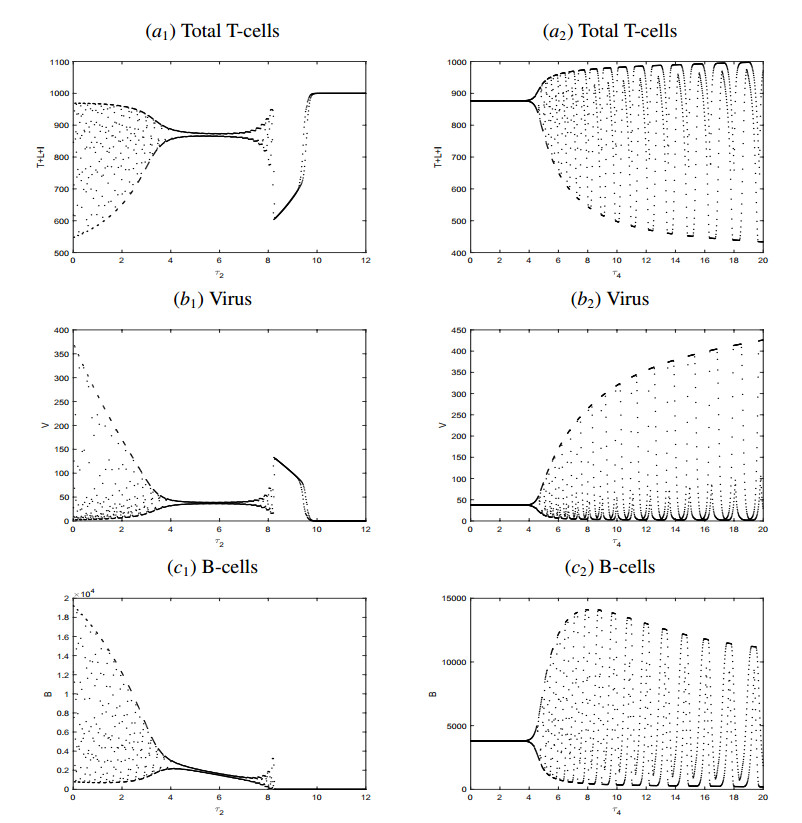
Figures(4) / Tables(1)

Yan Wang, Minmin Lu, Daqing Jiang. Viral dynamics of a latent HIV infection model with Beddington-DeAngelis incidence function, B-cell immune response and multiple delays[J]. Mathematical Biosciences and Engineering, 2021, 18(1): 274-299. doi: 10.3934/mbe.2021014







 DownLoad:
DownLoad: