Citation: Yanyan Du, Ting Kang, Qimin Zhang. Asymptotic behavior of a stochastic delayed avian influenza model with saturated incidence rate[J]. Mathematical Biosciences and Engineering, 2020, 17(5): 5341-5368. doi: 10.3934/mbe.2020289

| [1] | Centers for Disease Control and Prevention (CDC), Avian influenza, 2017. Available from: https://www.cdc.gov/flu/avianflu/influenza-a-virus-subtypes.htm. |
| [2] | S. Liu, S. Ruan, X. Zhang, Nonlinear dynamics of avian influenza epidemic models, Math. Biosci., 283 (2017), 118-135. |
| [3] | S. Iwami, Y. Takeuchi, X. Liu, Avian-human influenza epidemic model, Math. Biosci., 207 (2007), 1-25. |
| [4] | A. B. Gumel, Global dynamics of a two-strain avian influenza model, Int. J. Comput. Math., 86 (2009), 85-108. |
| [5] | Q. Tang, J. Ge, Z. Lin, An SEI-SI avian-human influenza model with diffusion and nonlocal delay, Appl. Math. Comput., 247 (2014), 753-761. |
| [6] | Y. Chen, Y. Wen, Global dynamic analysis of a H7N9 avian-human influenza model in an outbreak region, J. Theor. Biol., 367 (2015), 180-188. |
| [7] | S. Liu, S. Ruan, X. Zhang, On avian influenza epidemic models with time delay, Theor. Biosci., 134 (2015), 75-82. |
| [8] | N. S. Chong, J. M. Tchuenche, J. S. Robert, A mathematical model of avian influenza with halfsaturated incidence, Theor. Biosci., 133 (2013), 23-38. |
| [9] | Z. Liu, C. T. Fang, A modeling study of human infections with avian influenza A H7N9 virus in mainland China, Int. J. Infect. Dis., 41 (2015), 73-78. |
| [10] | C. Modnak, J. Wang, An avian influenza model with latency and vaccination, Dynam. Syst., 34 (2018), 195-217. |
| [11] | A. K. Muhammad, F. Muhammad, I. Saeed, B. Ebenezer, Modeling the transmission dynamics of avian influenza with saturation and psychological effect, Discrete Contin. Dyn. Syst. Ser. S, 12 (2019), 455-474. |
| [12] | T. Kang, Q. Zhang, L. Rong, A delayed avian influenza model with avian slaughter: Stability analysis and optimal control, Physica A, 529 (2019), 121544. |
| [13] | Y. Kuang, Delay Differential Equations with Applications in Population Dynamics, Academic Press, New York, 1993. |
| [14] | E. Beretta, Y. Takeuchi, Global stability of an sir epidemic model with time delays, J. Math. Biol., 33 (1995), 250-260. |
| [15] | M. Liu, C. Bai, K. Wang, Asymptotic stability of a two-group stochastic SEIR model with infinite delays, Commun. Nonlinear Sci. Numer. Simul., 19 (2014), 3444-3453. |
| [16] | Q. Liu, D. Jiang, N. Shi, T. Hayat, A. Alsaedi, Asymptotic behavior of a stochastic delayed seir epidemic model with nonlinear incidence, Physica A, 462 (2016), 870-882. |
| [17] | G. P. Samanta, Permanence and extinction for a nonautonomous avian-human influenza epidemic model with distributed time delay, Math. Comput. Model., 52 (2010), 1794-1811. |
| [18] | J. Semenza, B. Menne, Climate change and infectious diseases in europe, Lancet. Infect. Dis., 9 (2009), 365-375. |
| [19] | A. Lowen, J. Steel, Roles of humidity and temperature in shaping influenza seasonality, J. Virol., 88 (2014), 7692-7705. |
| [20] | X. Zhang, Global dynamics of a stochastic avian-human influenza epidemic model with logistic growth for avian population, Nonlinear Dynam., 90 (2017), 2331-2343. |
| [21] | F. Zhang, X. Zhang, The threshold of a stochastic avian-human influenza epidemic model with psychological effect, Physica A, 492 (2018), 485-495. |
| [22] | X. Zhang, Z. Shi, Y. Wang, Dynamics of a stochastic avian-human influenza epidemic model with mutation, Physica A, 534 (2019), 12940. |
| [23] | D. Jiang, J. Yu, C. Ji, N. Shi, Asymptotic behavior of global positive solution to a stochastic sir model, Math. Comput. Model., 54 (2011), 221-232. |
| [24] | Q. Liu, D. Jiang, T. Hayat, A. Alsaedi, Threshold behavior in a stochastic delayed sis epidemic model with vaccination and double diseases, J. Franklin I., 356 (2019), 7466-7485. |
| [25] | J. H. Desmond, An algorithmic introduction to numerical simulation of stochastic differential equations, SIAM Rev., 43 (2001), 525-546. |
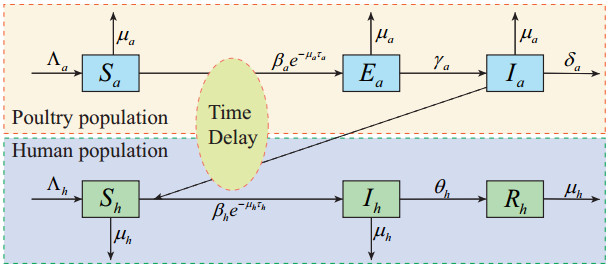
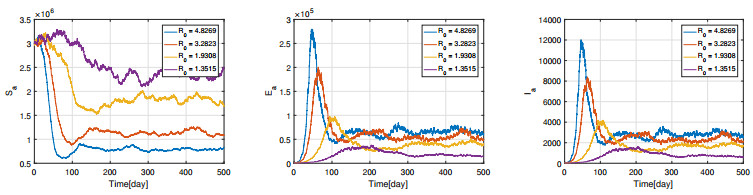
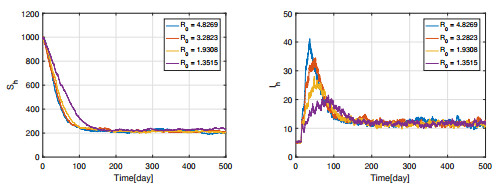
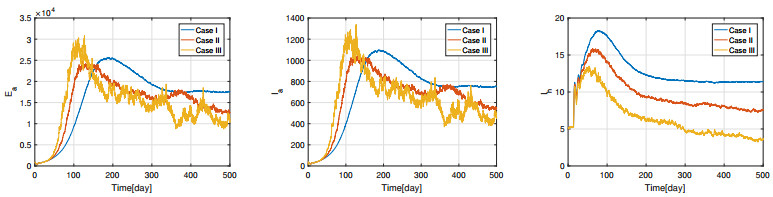
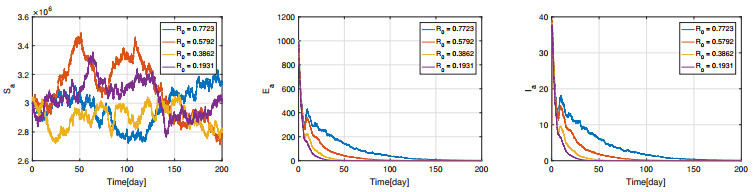
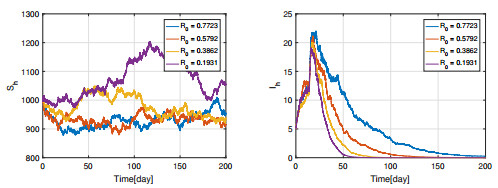
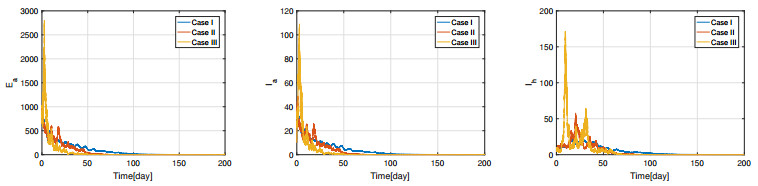
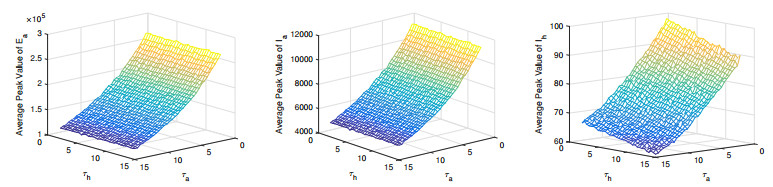
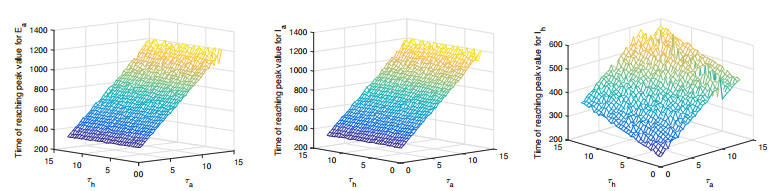
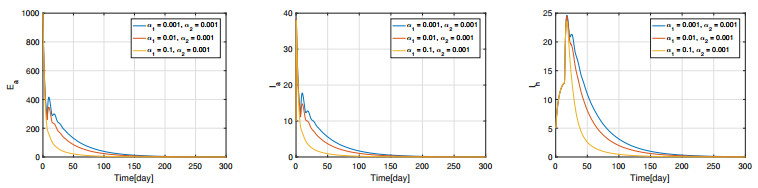
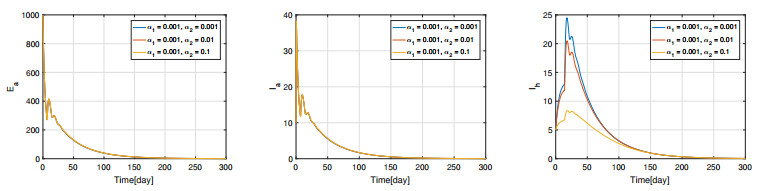
Figures(11) / Tables(3)

Yanyan Du, Ting Kang, Qimin Zhang. Asymptotic behavior of a stochastic delayed avian influenza model with saturated incidence rate[J]. Mathematical Biosciences and Engineering, 2020, 17(5): 5341-5368. doi: 10.3934/mbe.2020289







 DownLoad:
DownLoad: