Citation: Khalid Hattaf, Noura Yousfi. Dynamics of SARS-CoV-2 infection model with two modes of transmission and immune response[J]. Mathematical Biosciences and Engineering, 2020, 17(5): 5326-5340. doi: 10.3934/mbe.2020288

| [1] | A. E. Gorbalenya, S. C. Baker, R. S. Baric, R. J. de Groot, C. Drosten, A. A. Gulyaeva, et al., The species severe acute respiratory syndromerelated coronavirus: classifying 2019-nCoV and naming it SARS-CoV-2, Nat. Microbiol., 5 (2020), 536. |
| [2] | WHO, Coronavirus disease 2019 (COVID-19), Situation Report-139, 2020. Available from: https://www.who.int/docs/default-source/coronaviruse/situation-reports/20200607-covid-19-sitrep-139. |
| [3] | K. Hattaf, N. Yousfi, Qualitative analysis of a generalized virus dynamics model with both modes of transmission and distributed delays, Int. J. Differ. Equations, 2018 (2018), 1-7. |
| [4] | C. Li, J. Xu, J. Liu, Y. Zhou, The within-host viral kinetics of SARS-CoV-2, Math. Biosci. Eng., 17 (2020), 2853-2861. |
| [5] | M. A. Nowak, C. R. Bangham, Population dynamics of immune responses to persistent viruses, Science, 272 (1996), 74-79. |
| [6] | M. Dhar, S. Samaddar, P. Bhattacharya, R. K. Upadhyay, Viral dynamic model with cellular immune response: A case study of HIV-1 infected humanized mice, Physica A, 524 (2019), 1-14. |
| [7] | K. Hattaf, Spatiotemporal dynamics of a generalized viral infection model with distributed delays and CTL immune response, Computation, 7 (2019), 1-16. |
| [8] | H. R. Thieme, Persistence under relaxed point-dissipativity (with application to an endemic model), SIAM J. Math. Anal., 24 (1993), 407-435. |
| [9] | J. P. LaSalle, The Stability of Dynamical Systems, Regional Conference Series in Applied Mathematics, SIAM Philadelphia, 1976. |
| [10] | P. Roop-O, W. Chinviriyasit, S. Chinviriyasit, The effect of incidence function in backward bifurcation for malaria model with temporary immunity, Math. Biosci., 265 (2015), 47-64. |
| [11] | C. Yang, X. Wang, D. Gao and J. Wang, Impact of awareness programs on Cholera dynamics: Two modeling approaches, Bull. Math. Biol., 79 (2017), 2109-2131. |
| [12] | K. Hattaf, Global stability and Hopf bifurcation of a generalized viral infection model with multidelays and humoral immunity, Physica A, 545 (2020), 123689. |
| [13] | M. Ochs, J. R. Nyengaard, A. Jung, L. Knudsen, M. Voigt, T. Wahlers, J. Richter, H. G. Gundersen, The number of alveoli in the human lung, Am. J. Respir. Crit. Care Med., 169 (2004), 120-124. |
| [14] | P. Gehr, M. Bachofen, E. R. Weibel, The normal human lung: ultrastructure and morphometric estimation of diffusion capacity, Resp. Physiol., 32 (1978), 121-140. |
| [15] | J. D. Crapo, B. E. Barry, P. Gehr, M. Bachofen, E. R. Weibel, Cell number and cell characteristics of the normal human lung, Am. Rev. Respir. Dis., 126 (1982), 332-337. |
| [16] | E. R. Weibel, What makes a good lung? The morphometric basis of lung function, Swiss Med. Wkly., 139 (2009), 375-386. |
| [17] | H. Y. Lee, D. J. Topham, S. Y. Park, J. Hollenbaugh, J. Treanor, T. R Mosmann, et al., Simulation and prediction of the adaptive immune response to influenza A virus infection, J. Virol., 83 (2009), 7151-7165. |
| [18] | N. van Doremalen, T. Bushmaker, D. H. Morris, M. G. Holbrook, A. Gamble, B. N. Williamson, et al., Aerosol and surface stability of SARS-CoV-2 as compared with SARS-CoV-1, N. Engl. J. Med., 382 (2020), 1564-1567. |
| [19] | P. Czuppon, F. Débarre, A. Goncalves, O. Tenaillon, A. S. Perelson, J. Guedj, et al., Predicted success of prophylactic antiviral therapy to block or delay SARS-CoV-2 infection depends on the targeted mechanism, MedRxiv, (2020). |
| [20] | A. Gonçalves, J. Bertrand, R. Ke, E. Comets, X. de Lamballerie, D. Malvy, et al., Timing of antiviral treatment initiation is critical to reduce SARS-Cov-2 viral load, MedRxiv, (2020). |
| [21] | Y. M. Bar-On, A. Flamholz, R. Phillips, R. Milo, Science Forum: SARS-CoV-2 (COVID-19) by the numbers, Elife, 9 (2020), e57309. |
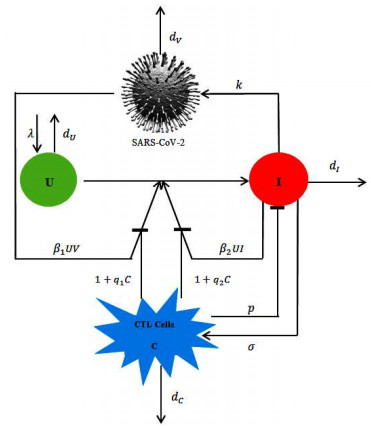
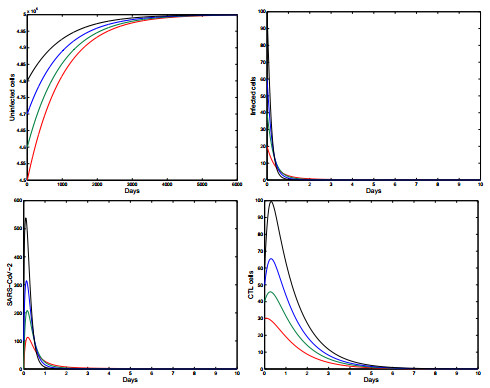
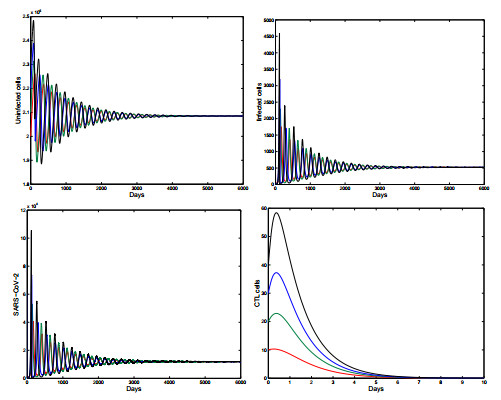
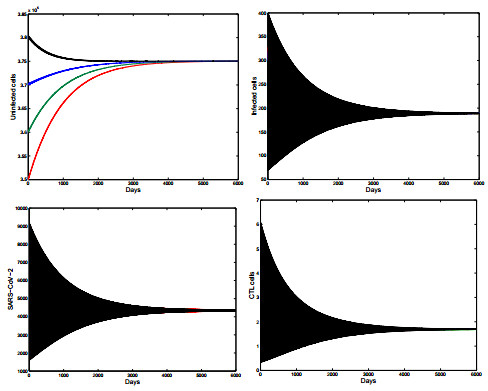
Figures(5) / Tables(1)

Khalid Hattaf, Noura Yousfi. Dynamics of SARS-CoV-2 infection model with two modes of transmission and immune response[J]. Mathematical Biosciences and Engineering, 2020, 17(5): 5326-5340. doi: 10.3934/mbe.2020288







 DownLoad:
DownLoad: