Citation: Farinaz Forouzannia, Sivabal Sivaloganathan, Mohammad Kohandel. A mathematical study of the impact of cell plasticity on tumour control probability[J]. Mathematical Biosciences and Engineering, 2020, 17(5): 5250-5266. doi: 10.3934/mbe.2020284

| [1] | D. Hanahan, R. A. Weinberg, Hallmarks of cancer: the next generation, Cell, 144 (2011), 646-674. |
| [2] | F. Pajonk, E. Vlashi, W. H. McBride, Radiation resistance of cancer stem cells: the 4 R's of radiobiology revisited, Stem Cells, 28 (2010), 639-648. |
| [3] | S. Sheila, C. Hawkins, et al, Identification of human brain tumour initiating cells, Nature, 432 (2004), 396-401. |
| [4] | N. D. Marjanovic, R. A. Weinberg, C. L. Chaffer, Poised with purpose: cell plasticity enhances tumorigenicity, Cell Cycle, 12 (2013), 2713-2714. |
| [5] | N. D. Marjanovic, R. A. Weinberg, C. L. Chaffer, Cell plasticity and heterogeneity in cancer, Clin. Chem., 59 (2013), 168-179. |
| [6] | F. Forouzannia, Studies of tumor heterogeneity, tumor microenvironment, and radiotherapy: A mathematical and computational approach, Ph.D thesis, University of Waterloo, 2019. |
| [7] | F. Forouzannia, H. Enderling, M. Kohandel, Mathematical Modeling of the Effects of Tumor Heterogeneity on the Efficiency of Radiation Treatment Schedule, Bull. Math. Biol., 80 (2018), 283-293. |
| [8] | F. Forouzannia, S. Sivaloganathan, Cancer Stem Cells, the Tipping Point: Minority Rules?, Curr. Stem Cell Rep., 3 (2017), 240-247. |
| [9] | R. Ganguly, I. K. Puri, Mathematical model for the cancer stem cell hypothesis, Cell Proliferation, 39 (2006), 3-14. |
| [10] | P. B. Gupta, C. M. Fillmore, G. Jiang, S. D. Shapira, K. Tao, C. Kuperwasser, et al., Stochastic State Transitions Give Rise to Phenotypic Equilibrium in Populations of Cancer Cells, Cell, 146 (2011), 633-644. |
| [11] | F. Michor, T. P. Hughes, Y. Iwasa, S. Branford, N. P. Shah, C. L. Sawyers, et al., Dynamics of chronic myeloid leukaemia, Nature, 435 (2005), 1267-1270. |
| [12] | C. Turner, M. Kohandel, Investigating the link between epithelial-mesenchymal transition and the cancer stem cell phenotype: a mathematical approach, J. Theor. Biol., 265 (2010), 329-335. |
| [13] | S. L. Weekes, B. Barker, S. Bober, K. Cisneros, J. Cline, A. Thompson, et al., A multicompartment mathematical model of cancer stem cell-driven tumor growth dynamics, Bull. Math. Biol., 76 (2014), 1762-1782. |
| [14] | S. Zapperi, C. A. M. La Porta, Do cancer cells undergo phenotypic switching? The case for imperfect cancer stem cell markers, Sci. Rep., 2 (2012), 1-7. |
| [15] | D. Zhou, Y. Wang, B. Wu, A multi-phenotypic cancer model with cell plasticity, J. Theor. Biol., 357 (2014), 35-45. |
| [16] | A. Dawson, T. Hillen, Derivation of the Tumour Control Probability (TCP) from a Cell Cycle Model, Comput. Math. Methods Med., 7 (2006), 121-141. |
| [17] | J. Gong, M. M. Dos Santos, C. Finlay, T. Hillen, Are more complicated tumour control probability models better?, Math. Med. Biol., 30 (2011), 1-19. |
| [18] | W. S. Kendal, Technical Report A closed-form description of tumour control with fractionated radiotherapy and repopulation, Int J Radiat Biol, 73 (1998), 207-210. |
| [19] | S. L. Tucker, H. D. Thames, M. G. Taylor, How well is the probability of tumor cure after fractionated irradiation described by Poisson statistics?, J. Theor. Biol., 124 (1990), 273-282. |
| [20] | A. Y. Yakovlev, Comments on the distribution of clonogens in irradiated tumors, Radiat. Res., 134 (1993), 117-120. |
| [21] | M. Zaider, L. Hanin, Tumor control probability in radiation treatment, Med. Phys., 38 (2011), 574-583. |
| [22] | M. Zaider, G. N. Minerbo, Tumour control probability: a formulation applicable to any temporal protocol of dose delivery, Phys. Med. Biol., 45 (2000), 279. |
| [23] | A. Dhawan, M. Kohandel, R. Hill, S. Sivaloganathan, Tumour control probability in cancer stem cells hypothesis, PloS one, 9 (2014), 121-141. |
| [24] | V. Shahrezaei, J. F. Ollivier, P. S. Swain, Colored extrinsic fluctuations and stochastic gene expression, Mol. Syst. Biol., 4 (2008). |
| [25] | E. J. Hall, A. J. Giaccia, Radiobiology for the Radiologist, 7th edition, Lippincott Williams Wilkins, Philadelphia, 2012. |
| [26] |
S. Bao, Q. Wu, R. E. McLendon, Y. Hao, Q. Shi, A. B. Hjelmeland, et al., Glioma Stem Cells Promote Radioresistance by Preferential Activation of the DNA Damage Response, Nature, 444 (2006), 756-760. doi: 10.1038/nature05236

|
| [27] | C. Lagadec, E. Vlashi, L. Della Donna, Y. Meng, C. Dekmezian, K. Kim, et al., Survival and selfrenewing capacity of breast cancer initiating cells during fractionated radiation treatment, Breast Cancer Res., 12 (2010), R13. |
| [28] | S. G. M. Piccirillo, B. A. Reynolds,N. Zanetti, G. Lamorte, E. Binda, G. Broggi, et al., Bone morphogenetic proteins inhibit the tumorigenic potential of human brain tumour-initiating cells, Nature, 444 (2006), 761-765. |
| [29] | T. M. Phillips, W. H. Mcbride, F. Pajonk, The response of CD24-/low/CD44+ breast cancerinitiating cells to radiation, J. Natl. Cancer Inst., 24 (2006), 1777-1785. |
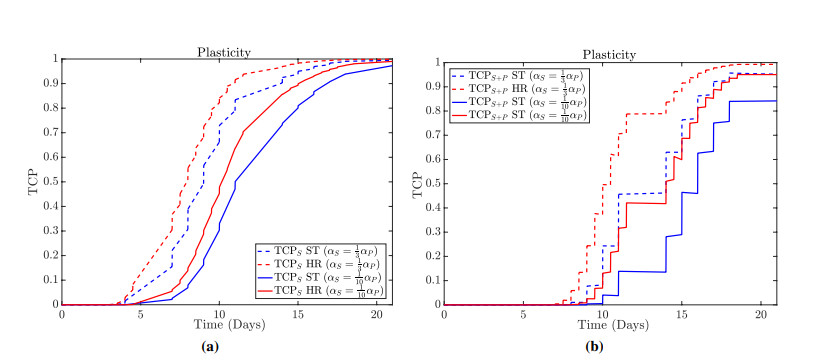
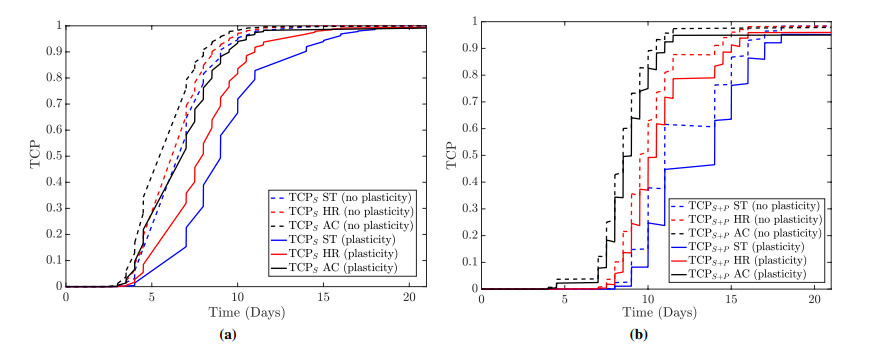
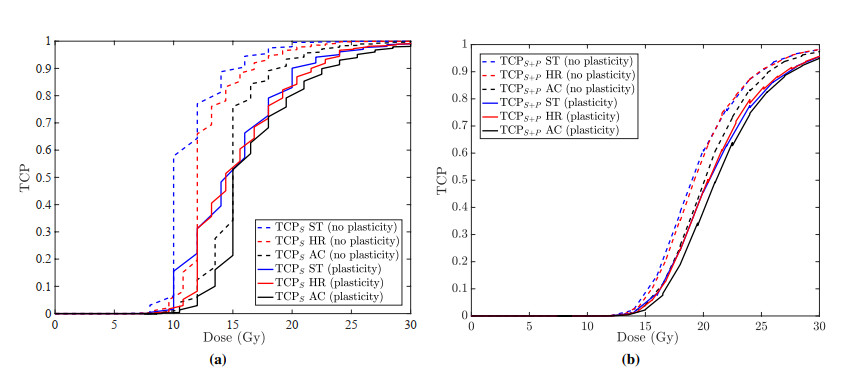
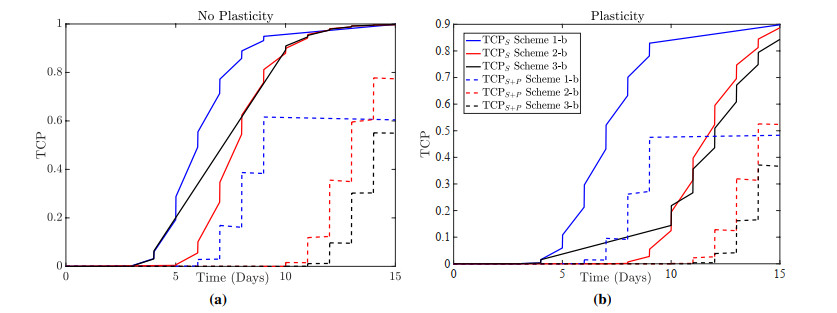
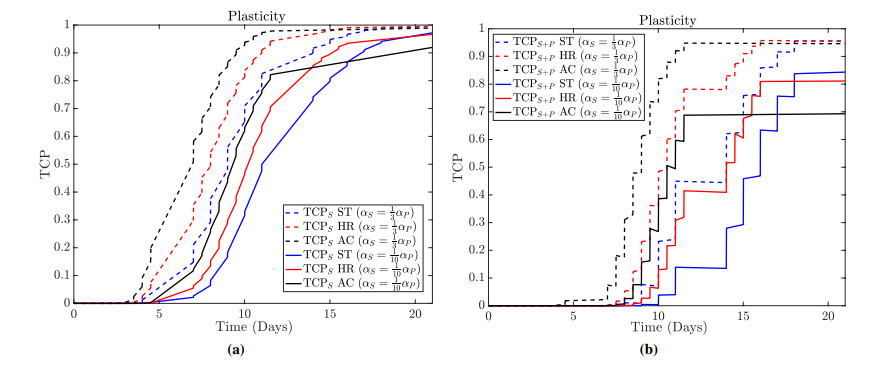
Figures(13)

Farinaz Forouzannia, Sivabal Sivaloganathan, Mohammad Kohandel. A mathematical study of the impact of cell plasticity on tumour control probability[J]. Mathematical Biosciences and Engineering, 2020, 17(5): 5250-5266. doi: 10.3934/mbe.2020284







 DownLoad:
DownLoad: