Citation: Philip A. Warrick, Emily F. Hamilton. Information theoretic measures of perinatal cardiotocography synchronization[J]. Mathematical Biosciences and Engineering, 2020, 17(3): 2179-2192. doi: 10.3934/mbe.2020116

| [1] | J. A. Low, R. Victory, E. J. Derrick, Predictive value of electronic fetal monitoring for intrapartum fetal asphyxia with metabolic acidosis, Obstet. Gynecol., 93 (1999), 285-291. |
| [2] | P. A. Warrick, E. F. Hamilton, D. Precup, R. Kearney, Classification of normal and hypoxic fetuses from systems modeling of intrapartum cardiotocography, IEEE Trans. Biomed. Eng., 57 (2010), 771-779. |
| [3] | M. Signorini, G. Magenes, S. Cerutti, D. Arduini, Linear and nonlinear parameters for the analysis of fetal heart rate signal from cardiotocographic recordings, IEEE Trans. Biomed. Eng., 50 (2003), 365-374. |
| [4] | F. Marzbanrad, Y. Kimura, M. Palaniswami, A. H. Khandoker, Quantifying the interactions between maternal and fetal heart rates by transfer entropy, PLoS ONE, 10 (2015), e0145672. |
| [5] | P. A. Warrick, E. F. Hamilton, Mutual information estimates of CTG synchronization, in Computing in Cardiology, 42 (2015), 137-139. |
| [6] | P. A. Warrick, E. F. Hamilton, D. Precup, R. E. Kearney, Identification of the dynamic relationship between intra-partum uterine pressure and fetal heart rate for normal and hypoxic fetuses, IEEE Trans. Biomed. Eng., 56 (2009), 1587-1597. |
| [7] | S. Haykin, Neural Networks: A Comprehensive Foundation, Prentice Hall, 1998. |
| [8] | P. Wollstadt, M. Martínez-Zarzuela, R. Vicente, F. J. Díaz-Pernas, M. Wibral, Efficient transfer entropy analysis of non-stationary neural time series, PLOS ONE, 9 (2014), 1-21. |
| [9] | J. Jezewski, K. Horoba, A. Matonia, J. Wrobel, Quantitative analysis of contraction patterns in electrical activity signal of pregnant uterus as an alternative to mechanical approach, Physiol. Meas., 26 (2005), 753. |
| [10] | P. Wollstadt, J. T. Lizier, R. Vicente, C. Finn, M. Martínez-Zarzuela, P. Mediano, et al., IDTxl: The Information Dynamics Toolkit xl: a Python package for the efficient analysis of multivariate information dynamics in networks, J. Open Source Software, 4 (2018), 1081. |
| [11] | A. Kraskov, H. Stögbauer, P. Grassberger, Estimating mutual information, Phys. Rev. E, 69 (2004), 066138, |
| [12] | L. Faes, G. Nollo, A. Porta, Information-based detection of nonlinear Granger causality in multivariate processes via a nonuniform embedding technique, Phys. Rev. E, 83 (2011), 051112. |
| [13] | T. Schreiber, Measuring information transfer, Phys. Rev. Lett., 85 (2000), 461-464. |
| [14] | M. Wibral, N. Pampu, V. Priesemann, F. Siebenhühner, H. Seiwert, M. Lindner, et al., Measuring information-transfer delays, PLoS ONE, 8 (2013), e55809. |
| [15] | M. Wibral, J. T. Lizier, S. Vögler, V. Priesemann, R. Galuske, Local active information storage as a tool to understand distributed neural information processing, Front. Neuroinf., 8 (2014), 1. |
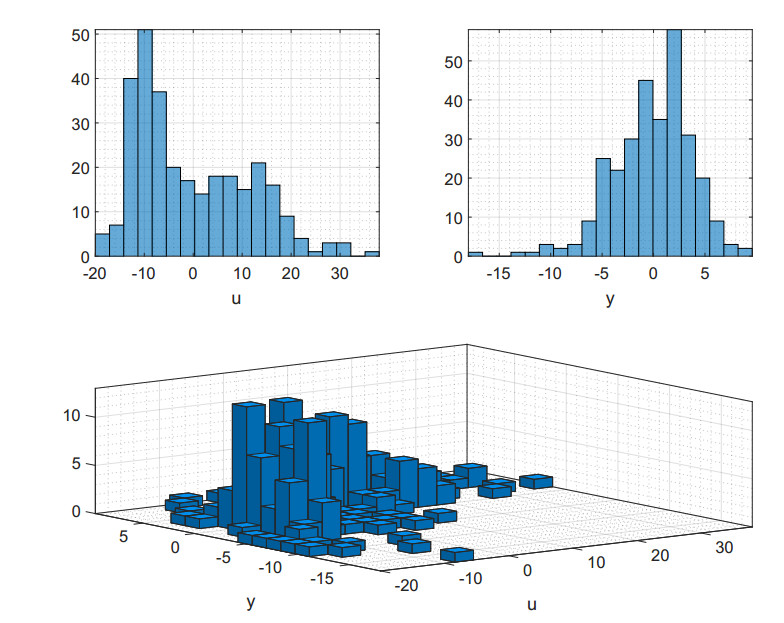
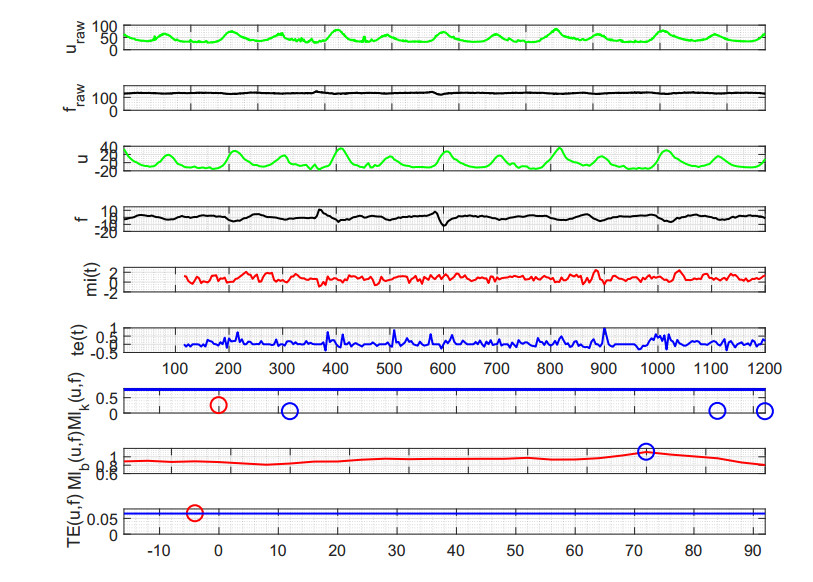
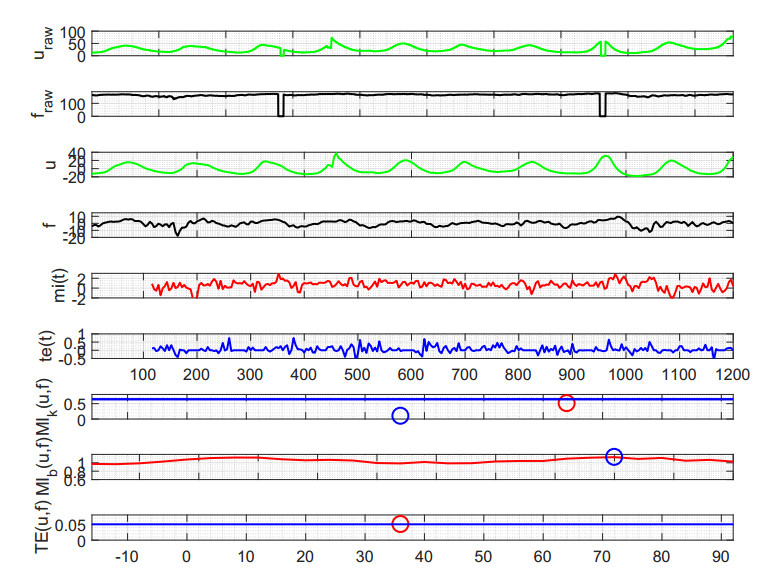
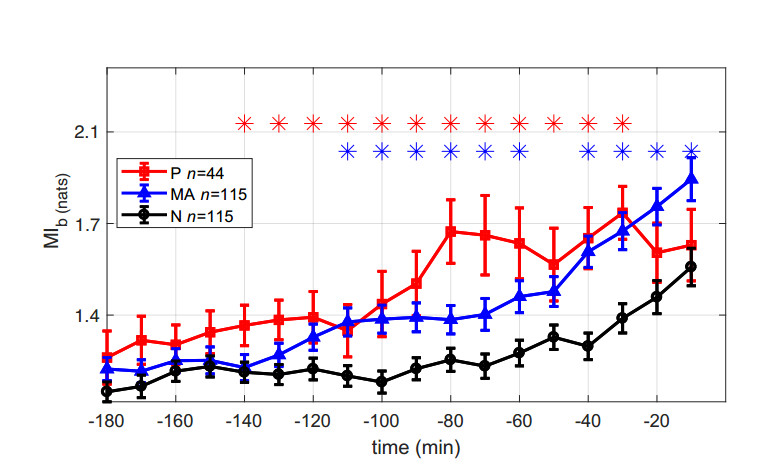
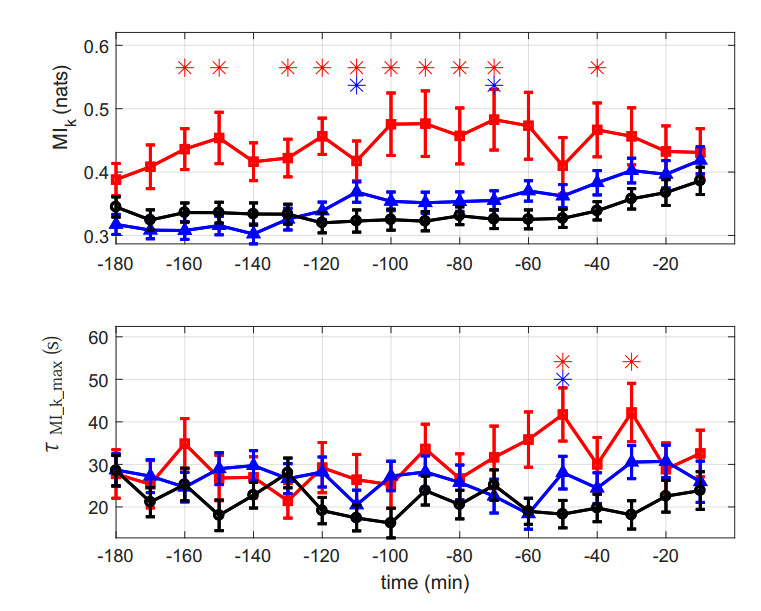
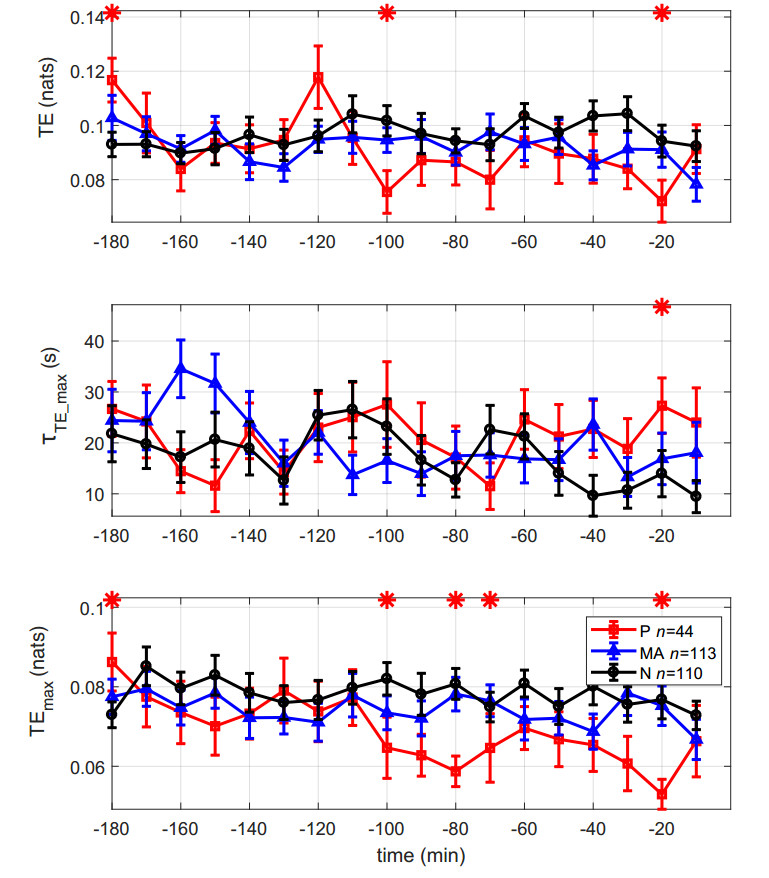
Figures(6) / Tables(1)

Philip A. Warrick, Emily F. Hamilton. Information theoretic measures of perinatal cardiotocography synchronization[J]. Mathematical Biosciences and Engineering, 2020, 17(3): 2179-2192. doi: 10.3934/mbe.2020116







 DownLoad:
DownLoad: