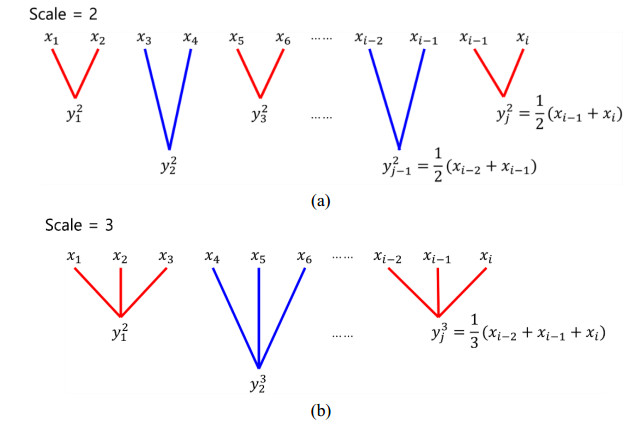
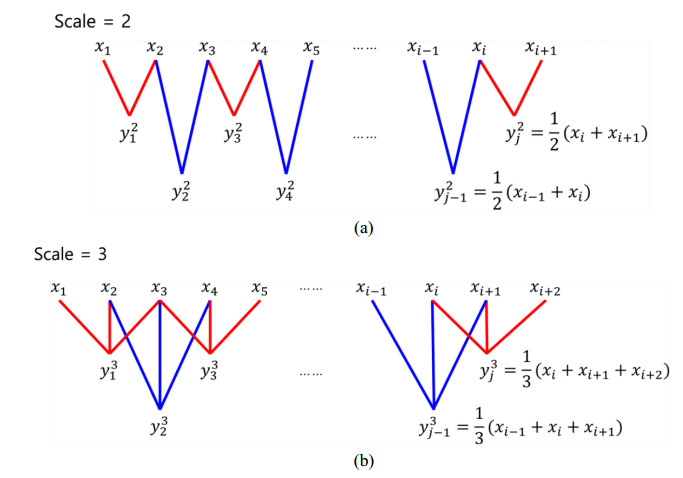
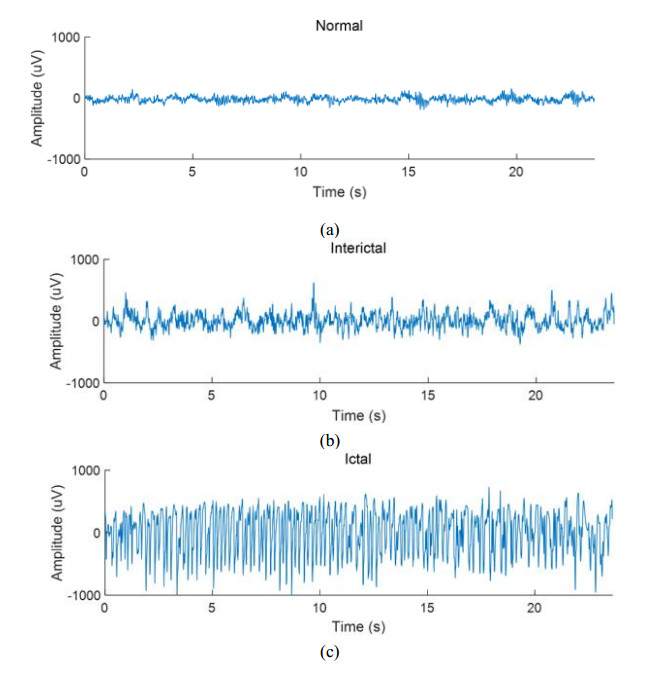
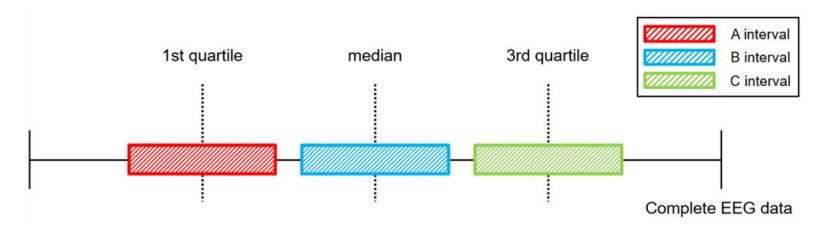
This paper proposes an information-theoretic measure for discriminating epileptic patterns in short-term electroencephalogram (EEG) recordings. Considering nonlinearity and nonstationarity in EEG signals, quantifying complexity has been preferred. To decipher abnormal epileptic EEGs, i.e., ictal and interictal EEGs, via short-term EEG recordings, a distribution entropy (DE) is used, motivated by its robustness on the signal length. In addition, to reflect the dynamic complexity inherent in EEGs, a multiscale entropy analysis is incorporated. Here, two multiscale distribution entropy (MDE) methods using the coarse-graining and moving-average procedures are presented. Using two popular epileptic EEG datasets, i.e., the Bonn and the Bern-Barcelona datasets, the performance of the proposed MDEs is verified. Experimental results show that the proposed MDEs are robust to the length of EEGs, thus reflecting complexity over multiple time scales. In addition, the proposed MDEs are consistent irrespective of the selection of short-term EEGs from the entire EEG recording. By evaluating the Man-Whitney U test and classification performance, the proposed MDEs can better discriminate epileptic EEGs than the existing methods. Moreover, the proposed MDE with the moving-average procedure performs marginally better than one with the coarse-graining. The experimental results suggest that the proposed MDEs are applicable to practical seizure detection applications.
Citation: Dae Hyeon Kim, Jin-Oh Park, Dae-Young Lee, Young-Seok Choi. Multiscale distribution entropy analysis of short epileptic EEG signals[J]. Mathematical Biosciences and Engineering, 2024, 21(4): 5556-5576. doi: 10.3934/mbe.2024245
This paper proposes an information-theoretic measure for discriminating epileptic patterns in short-term electroencephalogram (EEG) recordings. Considering nonlinearity and nonstationarity in EEG signals, quantifying complexity has been preferred. To decipher abnormal epileptic EEGs, i.e., ictal and interictal EEGs, via short-term EEG recordings, a distribution entropy (DE) is used, motivated by its robustness on the signal length. In addition, to reflect the dynamic complexity inherent in EEGs, a multiscale entropy analysis is incorporated. Here, two multiscale distribution entropy (MDE) methods using the coarse-graining and moving-average procedures are presented. Using two popular epileptic EEG datasets, i.e., the Bonn and the Bern-Barcelona datasets, the performance of the proposed MDEs is verified. Experimental results show that the proposed MDEs are robust to the length of EEGs, thus reflecting complexity over multiple time scales. In addition, the proposed MDEs are consistent irrespective of the selection of short-term EEGs from the entire EEG recording. By evaluating the Man-Whitney U test and classification performance, the proposed MDEs can better discriminate epileptic EEGs than the existing methods. Moreover, the proposed MDE with the moving-average procedure performs marginally better than one with the coarse-graining. The experimental results suggest that the proposed MDEs are applicable to practical seizure detection applications.

| [1] |
A. K. Ngugi, C. Bottomley, I. Kleinschmidt, J. W. Sander, C. R. Newton, Estimation of the burden of active and life-time epilepsy: A meta-analytic approach, Epilepsia, 51 (2010), 883–890. https://doi.org/10.1111/j.1528-1167.2009.02481.x doi: 10.1111/j.1528-1167.2009.02481.x

|
| [2] | W. A. Hauser, E. Beghi, First seizure definitions and worldwide incidence and mortality, Epilepsia, 49 (2008), 8–12. |
| [3] |
R. S. Fisher, C. Acevedo, A. Arzimanoglou, A. Bogacz, J. H. Cross, C. E. Elger, et al., ILAE official report: A practical clinical definition of epilepsy, Epilepsia, 55 (2014), 475–482. https://doi.org/10.1111/epi.12550 doi: 10.1111/epi.12550
|
| [4] | O. Devinsky, T. Spruill, D. Thurman, D. Friedman, Recognizing and preventing epilepsy-related mortality: A call for action, Neurology, 86 (2016), 779–786. |
| [5] |
H. O. Lüders, I. Najm, D. Nair, P. Widdess-Walsh, W. Bingman, The epileptogenic zone: general principles, Epileptic Disord., 8 (2006), 1–2. https://doi.org/10.1684/j.1950-6945.2006.tb00152.x doi: 10.1684/j.1950-6945.2006.tb00152.x
|
| [6] |
Y. Paul, Various epileptic seizure detection techniques using biomedical signals: A review, Brain Inf., 5 (2018). https://doi.org/10.1186/s40708-018-0084-z doi: 10.1186/s40708-018-0084-z
|
| [7] |
S. Noachtar, J. Rémi, The role of EEG in epilepsy: A critical review, Epilepsy Behav., 15 (2009), 22–33. https://doi.org/10.1016/j.yebeh.2009.02.035 doi: 10.1016/j.yebeh.2009.02.035
|
| [8] |
U. Seneviratne, M. Cook, W. D'Souza, Brainwaves beyond diagnosis: Wider applications of electroencephalography in idiopathic generalized epilepsy, Epilepsia, 63 (2022), 22–41. https://doi.org/10.1111/epi.17119 doi: 10.1111/epi.17119
|
| [9] |
R. Cherian, E. G. Kanaga, Theoretical and methodological analysis of EEG based seizure detection and prediction: An exhaustive review, J. Neurosci. Methods, 369 (2022), 109483. https://doi.org/10.1016/j.jneumeth.2022.109483 doi: 10.1016/j.jneumeth.2022.109483
|
| [10] |
S. J. J. Jui, R. C. Deo, P. D. Barua, A. Devi, J. Soar, U. R. Acharya, Application of entropy for automated detection of neurological disorders with electroencephalogram signals: A review of the last decade (2012–2022), IEEE Access, 11 (2023), 71905–71924. https://doi.org/10.1109/ACCESS.2023.3294473 doi: 10.1109/ACCESS.2023.3294473
|
| [11] |
A. Ulate-Campos, F. Coughlin, M. Gaínza-Lein, I. S. Fernández, P. L. Pearl, T. Loddenkemper, Automated seizure detection systems and their effectiveness for each type of seizure, Seizure, 40 (2016), 88–101. https://doi.org/10.1016/j.seizure.2016.06.008 doi: 10.1016/j.seizure.2016.06.008
|
| [12] |
A. Shoeibi, N. Ghassemi, R. Alizadehsani, M. Rouhani, H. Hosseini-Nejad, A. Khosravi, et al., A comprehensive comparison of handcrafted features and convolutional autoencoders for epileptic seizures detection in EEG signals, Expert Syst. Appl., 163 (2021), 113788. https://doi.org/10.1109/ACCESS.2023.3294473 doi: 10.1109/ACCESS.2023.3294473
|
| [13] |
D. Y. Lee, Y. S. Choi, Multiscale distribution entropy analysis of short-term heart rate variability, Entropy, 20 (2018), 952. https://doi.org/10.3390/e20120952 doi: 10.3390/e20120952
|
| [14] |
V. Srinivasan, C. Eswaran, N. Sriraam, Approximate entropy-based epileptic EEG detection using artificial neural networks, IEEE Trans. Inf. Technol. Biomed., 11 (2007), 288–295. https://doi.org/10.1109/TITB.2006.884369 doi: 10.1109/TITB.2006.884369
|
| [15] |
H. Ocak, Automatic detection of epileptic seizures in EEG using discrete wavelet transform and approximate entropy, Expert Syst. Appl., 36 (2009), 2027–2036. https://doi.org/10.1016/j.eswa.2007.12.065 doi: 10.1016/j.eswa.2007.12.065
|
| [16] |
J. S. Richman, J. R. Moorman, Physiological time-series analysis using approximate entropy and sample entropy, Am. J. Physiol. Heart Circ. Physiol., 278 (2000), H2039–2049. https://doi.org/10.1152/ajpheart.2000.278.6.H2039 doi: 10.1152/ajpheart.2000.278.6.H2039
|
| [17] |
Y. Kumar, M. L. Dewal, R. S. Anand, Epileptic seizure detection using DWT based fuzzy approximate entropy and support vector machine, Neurocomputing, 133 (2014), 271–279. https://doi.org/10.1016/j.neucom.2013.11.009 doi: 10.1016/j.neucom.2013.11.009
|
| [18] |
Y. S. Choi, K. Hyun, J. Y. Choi, Assessing multiscale permutation entropy for short electroencephalogram recordings, Cluster Comput., 19 (2016), 2305–2314. https://doi.org/10.1007/s10586-016-0648-8 doi: 10.1007/s10586-016-0648-8
|
| [19] |
M. Costa, A. L. Goldberger, C. K. Peng, Multiscale entropy analysis of complex physiologic time series, Phys. Rev. Lett., 89 (2002), 068102. https://doi.org/10.1103/PhysRevLett.89.068102 doi: 10.1103/PhysRevLett.89.068102
|
| [20] |
S. D. Wu, C. W. Wu, K. Y. Lee, S. G. Lin, Modified multiscale entropy for short-term time series analysis, Phys. A, 392 (2013), 5865–5873. https://doi.org/10.1016/j.physa.2013.07.075 doi: 10.1016/j.physa.2013.07.075
|
| [21] |
Y. Zhang, P. Shang, Refined composite multiscale weighted-permutation entropy of financial time series, Phys. A, 496 (2018), 189–199. https://doi.org/10.1016/j.physa.2013.07.075 doi: 10.1016/j.physa.2013.07.075
|
| [22] |
Y. Li, B. Tang, S. Jiao, Q. Su, Snake optimization-based variable-step multiscale single threshold slope entropy for complexity analysis of signals, IEEE Trans. Instrum. Meas., 72 (2023), 6505313. https://doi.org/10.1109/TIM.2023.3317908 doi: 10.1109/TIM.2023.3317908
|
| [23] |
Y. Li, B. Tang, S. Jiao, Y. Zhou, Optimized multivariate multiscale slope entropy for nonlinear dynamic analysis of mechanical signals, Chaos Solitons Fractals, 179 (2024), 114436. https://doi.org/10.1016/j.chaos.2023.114436 doi: 10.1016/j.chaos.2023.114436
|
| [24] |
P. Li, C. Liu, K. Li, D. Zheng, C. Liu, Y. Hou, Assessing the complexity of short-term heartbeat interval series by distribution entropy, Med. Biol. Eng. Comput., 53 (2015), 77–87. https://doi.org/10.1016/j.chaos.2023.114436 doi: 10.1016/j.chaos.2023.114436
|
| [25] | R. K. Udhayakumar, C. Karmakar, P. Li, M. Palaniswami, Effect of data length and bin numbers on distribution entropy (DistEn) measurement in analyzing healthy aging, in 2015 37th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC), (2015), 7877–7880. https://doi.org/10.1109/EMBC.2015.7320218 |
| [26] |
P. Li, C. Karmakar, C. Yan, M. Palaniswami, C. Liu, Classification of 5-S Epileptic EEG recordings using distribution entropy and sample entropy, Front. Physiol., 7 (2016), 136. https://doi.org/10.3389/fphys.2016.00136 doi: 10.3389/fphys.2016.00136
|
| [27] |
D. Y. Lee, Y. S. Choi, Multiscale distribution entropy analysis of heart rate variability using differential inter-beat intervals, IEEE Access, 8 (2020), 48761–48773. https://doi.org/10.1109/ACCESS.2020.2978930 doi: 10.1109/ACCESS.2020.2978930
|
| [28] |
S. Supriya, S. Siuly, H. Wang, Y. Zhang, Epilepsy detection from eeg using complex network techniques: A Review, IEEE Rev. Biomed. Eng., 16 (2023), 292–306. https://doi.org/10.1109/RBME.2021.3055956 doi: 10.1109/RBME.2021.3055956
|
| [29] |
É. Lemoine, J. N. Briard, B. Rioux, R. Podbielski, B. Nauche, D. Toffa, et al., Computer-assisted analysis of routine electroencephalogram to identify hidden biomarkers of epilepsy: Protocol for a systematic review, BMJ Open, 13 (2023), e066932. https://doi.org/10.1136/bmjopen-2022-066932 doi: 10.1136/bmjopen-2022-066932
|
| [30] |
R. G. Andrzejak, K. Lehnertz, F. Mormann, C. Rieke, P. David, C. E. Elger, Indications of nonlinear deterministic and finite-dimensional structures in time series of brain electrical activity: Dependence on recording region and brain state, Phys. Rev. E, 64 (2001), 061907. https://doi.org/10.1103/PhysRevE.64.061907 doi: 10.1103/PhysRevE.64.061907
|
| [31] |
R. G. Andrzejak, K. Schindler, C. Rummel, Nonrandomness, nonlinear dependence, and nonstationarity of electroencephalographic recordings from epilepsy patients, Phys. Rev. E, 86 (2012), 046206. https://doi.org/10.1103/PhysRevE.86.046206 doi: 10.1103/PhysRevE.86.046206
|
| [32] | T. Gautama, D. P. Mandic, M. M. Van Hulle, A differential entropy based method for determining the optimal embedding parameters of a signal, in 2003 IEEE International Conference on Acoustics, Speech, and Signal Processing, (2023). |
| [33] |
M. R. Islam, X. Zhao, Y. Miao, H. Sugano, T. Tanaka, Epileptic seizure focus detection from interictal electroencephalogram: A survey, Cogn. Neurodyn., 17 (2023), 1–23. https://doi.org/10.1007/s11571-022-09816-z doi: 10.1007/s11571-022-09816-z
|
| [34] |
L. Ilias, D. Askounis, J. Psarras, Multimodal detection of epilepsy with deep neural networks, Expert Syst. Appl., 213 (2023), 119010. https://doi.org/10.1016/j.eswa.2022.119010 doi: 10.1016/j.eswa.2022.119010
|
| [35] |
M. Sunkara, S. R. Reeja, Tri-SeizureDualNet: A novel multimodal brain seizure detection using triple stream skipped feature extraction module entrenched dual parallel attention transformer, Biomed. Signal Process Control, 88 (2024), 105593. https://doi.org/10.1016/j.bspc.2023.105593 doi: 10.1016/j.bspc.2023.105593
|
| [36] |
F. Dong, Z. Yuan, D. Wu, L. Jiang, J. Liu, W. Hu, Novel seizure detection algorithm based on multi-dimension feature selection, Biomed. Signal Process Control, 84 (2023), 104747. https://doi.org/10.1016/j.bspc.2023.104747 doi: 10.1016/j.bspc.2023.104747
|
| [37] |
L. Jiang, J. He, H. Pan, D. Wu, T. Jiang, J. Liu, Seizure detection algorithm based on improved functional brain network structure feature extraction, Biomed. Signal Process Control, 79 (2023), 104053. https://doi.org/10.1016/j.bspc.2022.104053 doi: 10.1016/j.bspc.2022.104053
|
| [38] |
A. Einizade, S. Nasiri, M. Mozafari, S. H. Sardouie, G.D. Clifford, Explainable automated seizure detection using attentive deep multi-view networks, Biomed. Signal Process Control, 79 (2023), 104076. https://doi.org/10.1016/j.bspc.2022.104076 doi: 10.1016/j.bspc.2022.104076
|
| [39] |
X. Qiu, F. Yan, H. Liu, A difference attention ResNet-LSTM network for epileptic seizure detection using EEG signal, Biomed. Signal Process Control, 83 (2023), 104652. https://doi.org/10.1016/j.bspc.2023.104652 doi: 10.1016/j.bspc.2023.104652
|
| [40] |
D. Lee, B. Kim, T. Kim, I. Joe, J. Chong, K. Min, K. Jung, A ResNet-LSTM hybrid model for predicting epileptic seizures using a pretrained model with supervised contrastive learning, Sci. Rep., 14 (2024), 1319. https://doi.org/10.1038/s41598-023-43328-y doi: 10.1038/s41598-023-43328-y
|
| [41] | T. K. K. Ho, N. Armanfard, Self-supervised learning for anomalous channel detection in EEG graphs: application to seizure analysis, in Proceedings of the AAAI Conference on Artificial Intelligence, (2023), 7866–7874. https://doi.org/10.1609/aaai.v37i7.25952 |
Figures(8) / Tables(3)

Dae Hyeon Kim, Jin-Oh Park, Dae-Young Lee, Young-Seok Choi. Multiscale distribution entropy analysis of short epileptic EEG signals[J]. Mathematical Biosciences and Engineering, 2024, 21(4): 5556-5576. doi: 10.3934/mbe.2024245







 DownLoad:
DownLoad: