Citation: P. E. Greenwood, L. M. Ward. Rapidly forming, slowly evolving, spatial patterns from quasi-cycle Mexican Hat coupling[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 6769-6793. doi: 10.3934/mbe.2019338

| [1] | Y. Kuramoto, Chemical Oscillations, Waves and Turbulence, Springer, New York, 1984. |
| [2] | P. E. Greenwood, M. D. McDonnell and L. M. Ward, A Kuramoto coupling of quasi-cycle oscillators with application to neural networks, J. Coupl. Syst. Multiscale Dynam., 4, (2016), 1–12. |
| [3] | S. H. Strogatz, From Kuramoto to Crawford: Exploring the onset of synchronization in popula-tions of coupled oscillators, Physica D, 143, (2000), 1–20. |
| [4] | M. Breakspear, S. Heitmann and A. Daffertshofer, Generative models of cortical oscillations: neurobiological implications of the Kuramoto model, Front. Hum. Neurosci., 4, (2010), 190. |
| [5] | G. Deco and M. L. Kringelbach, Metastability and coherence: Extending the communication through coherence hypothesis using a whole-brain computational perspective, Trends Neurosci., 39, (2016), 125–135. |
| [6] | S. Chandra and M. Girvan and E. Ott, Complexity reduction for systems of interacting orientable agents: beyond the Kuramoto model, arXiv,1809.00783v1, (2018). |
| [7] | S. Heitman and G. B. Ermentrout, Synchrony, waves and ripple in spatially coupled Kuramoto oscillators with Mexican hat connectivity, Biol. Cybernet., 109, (2015), 333–347. |
| [8] | Y. Park, S. Heitman and G.B. Ermentrout, The utility of phase models in studying neural synchro-nization, arXiv, 1707.05713, (2017). |
| [9] | T. Uezu, T. Kimoto and M. Okada, Solvable model of a phase oscillator Network on a circle with infinite-range Mexican-Hat-type Interaction, J. Phys. Soc. Jap., 81, (2012), 07301. |
| [10] | T. Uezu, T. Kimoto and M. Okada, Analysis of a solvable model of a phase oscillator network on a circle with infinite-range Mexican-hat-type interaction, Phys. Rev. E, 88, (2013), 032918. |
| [11] | P. C. Bressloff, Metastable states and quasicycles in a stochastic Wilson-Cowan model of neuronal population dynamics, Phys. Rev. E, 82, (2010), 051903. |
| [12] | P. E. Greenwood, M. D. McDonnell and L. M. Ward, Dynamics of gamma bursts in local field potentials, Neural Comput., 27, (2015), 74–103. |
| [13] | L. M. Ward and P. E. Greenwood, Stochastic facilitation in the brain? J. Stat. Mech. Theory Exper., 5, (2016), 054033. |
| [14] | G. von Bekesy, Mach band type lateral inhibition in different sense organs, J. Gen. Physiol., 50, (1967), 519–532. |
| [15] | D. Marr, Vision, Academic Press, New York, 1980. |
| [16] | K. Hamaguchi and K. Aihara, Quantitative information transfer through layers of spiking neurons connected by Mexican-Hat-type connectivity, Neurocomputing., 58–60, (2004), 85–90. |
| [17] | S. P. Burns, D. Xing and R. M. Shapley, Is gamma-band activity in the local field potential of V1 cortex a "clock" or filtered noise? J. Neurosci., 31, (2011), 9658–9664. |
| [18] | T. C. Butler and N. Goldenfeld, Fluctuation-driven Turing patterns, Phys. Rev. E, 84, (2011), 011112. |
| [19] | T. C. Butler, M. Benayoun, E. Wallace, et al., Evolutionary constraints on visual cortex architecture from the dynamics of hallucinations, Proc. Nat. Acad. Sci. USA, 109, (2012), 606–609. |
| [20] | A. J. McKane, T. Biancalani and T. Rogers, Stochastic pattern formation and spontaneous polari-sation: The linear noise approximation and beyond, Bull. Math. Biol., 76, (2014), 895–921. |
| [21] | P. H. Baxendale, P. E. Greenwood and L. M. Ward, Noise sharing and Mexican Hat coupling in a stochastic neural field, Phys. Rev. E, (2019). |
| [22] | P. H. Baxendale and P. E. Greenwood, Sustained oscillations for density dependent Markov processes, J. Math. Biol., 63, (2011), 433–457. |
| [23] | K. Kang, M. Shelley, J. A. Henrie, et al., LFP spectral peaks in V1 cortex: Network resonance and cortico-cortical feedback, J. Computat. Neurosci., 29, (2010), 495–507. |
| [24] | S. L. Brincat and C. E. Connor, Dynamic shape synthesis in posterior inferotemporal cortex, Neuron, 49, (2006), 17–24. |
| [25] | E. Salinas and L. F. Abbott, A model of multiplicative neural responses in parietal cortex, Proc. Nat. Acad. Sci. USA, 93, (1996), 11956–11961. |
| [26] | R. E. Walley and T. D. Weiden, Lateral inhibition and cognitive masking: A neuropsychological theory of attention, Psych. Rev., 80, (1973), 284–302. |
| [27] | K. Kang, M. Shelley and H. Sompolinsky, Mexican hats and pinwheels in visual cortex, Proc. Nat. Acad. Sci. USA, 100, (2003), 2848–2853. |
| [28] | P. Buzás, U. T. Eysel, P. Adjorán, et al., Axonal topography of cortical basket cells in relation to orientation, direction, and ocular dominance maps, J. Comp. Neurol., 437, (2001), 259–285. |
| [29] | J. P. Lachaux, E. Rodriguez, J. Martinerie, et al., Measuring phase synchrony in brain signals, Hum. Brain Map., 8, (1999), 194–208. |
| [30] | P. Fries, A mechanism for cognitive dynamics: Neuronal communication through neuronal coher- ence, Trends Cog. Sci., 9, (2005), 474–480. |
| [31] | H. P. Wang, D. Spencer, J. M. Fellous, et al., Synchrony of thalamocortical inputs maximizes cortical reliability, Science, 328, (2010), 106–109. |
| [32] | P. E. Kloeden and E. Platen, Numerical Solution of Stochastic Differential Equations, Springer-Verlag, New York, 1992. |
| [33] | J. D. Murray, Mathematical Biology, Springer, Berlin, 1989. |
| [34] | B. van der Pol, On "relaxation-oscillations", Phil. Mag. J. Sci., 2, (1926), 978–992. |
| [35] | D. Somers and N. Kopell, Rapid synchrony through fast threshold modulation, Biol. Cyber., 68, (1993), 393–407. |
| [36] | A. Daffertshofer and B. C. M.van Wijk, On the influence of amplitude on the connectivity between phases, Frontiers Neuroinform., 5, (2011), 1–12. |
| [37] | T. Akam and D. M. Kullmann, Oscillatory multiplexing of population codes for selective com-munication in the mammalian brain, Nat. Rev. Neurosci., 15, (2014), 111–122. |
| [38] | A. L. Barth and J. F. A. Poulet, Experimental evidence for sparse firing in the neocortex, Trends Neurosci., 35, (2012), 345–355. |
| [39] | E. Wallace, M. Benayoun, W. van Dronglen, et al., Emergent oscillations in networks of stochastic spiking neurons, PLoS One, 6, (2011), e14804. |
| [40] | D. Terman and D Wang, Global competition and local cooperation in a network of neural oscilla-tors, Physica D, 81, (1995), 148–176. |
| [41] | O. Faugeras and J. Inglis, Stochastic neural field equations: a rigorous footing, J. Math. Biol., 71, (2015), 259–300. |
| [42] | A. Hutt, A. Longtin and L. Schimansky-Geier, Additive noise-induced Turing transitions in spatial systems with application to neural fields and the Swift-Hohenberg equation, Physica D, 237, (2008), 755–773. |
| [43] | P. Jung and G. Mayer-Kress Spatiotemporal stochastic resonance in excitable media Phys. Rev. Lett., 74 ,(1995), 2130–2133. |
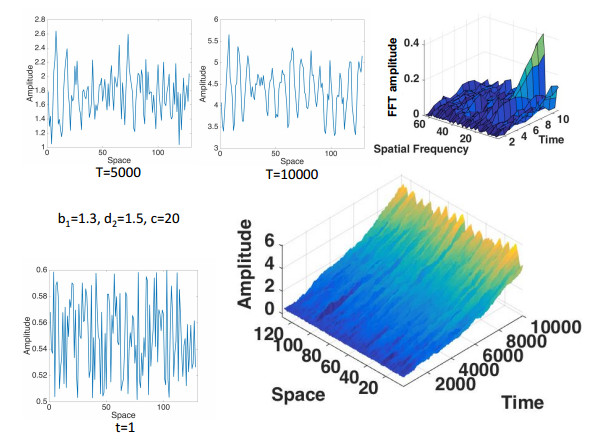
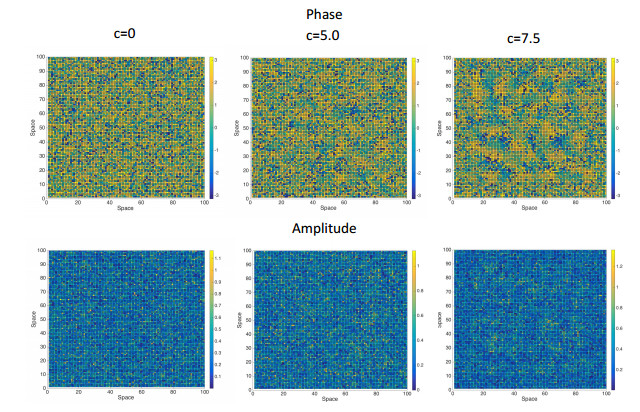
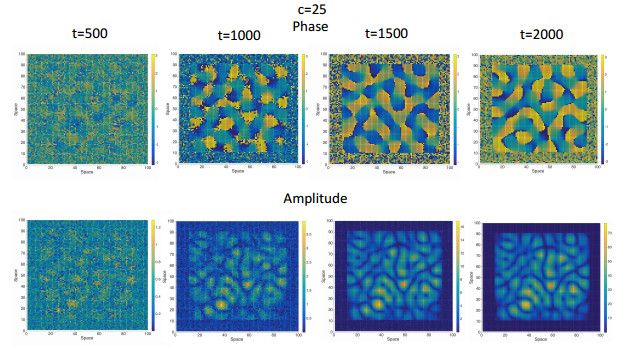
Figures(7) / Tables(1)

P. E. Greenwood, L. M. Ward. Rapidly forming, slowly evolving, spatial patterns from quasi-cycle Mexican Hat coupling[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 6769-6793. doi: 10.3934/mbe.2019338







 DownLoad:
DownLoad: