Citation: Yunbo Tu, Shujing Gao, Yujiang Liu, Di Chen, Yan Xu. Transmission dynamics and optimal control of stage-structured HLB model[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 5180-5205. doi: 10.3934/mbe.2019259

| [1] | D. Farnsworth, K. A. Grogan, A. H. C. V. Bruggen, et al., The potential economic cost and response to greening in Florida citrus, Agric. Appl. Econ. Assoc., 29 (2014): 1–6. |
| [2] | J. M. Bove, Huanglongbing: A destructive, newly-emerging, century-old disease of citrus, J. Plant. Pathol., 88 (2006): 7–37. |
| [3] | S. Alvarez, D. Solis and M. Thomas, Can Florida's citrus industry be saved while preserving the environment? An economic analysis for the bio-control of the Asian Citrus Psyllid, S. Agric. Econ. Assoc., 2015. |
| [4] | H. Su, Research and health management of citrus Huanglongbing in Taiwan, In: Proc Intern Research Conference on Huanglongbing, Orlando, Florida, USA, (2008): 57–92. |
| [5] | A. J. Ayres, J. B. Jr and J. M. Bové, The experience with Huanglongbing management in Brazil, Acta. Hortic., 1065 (2015): 55–62. |
| [6] | W. Li, Y. N. Yao, L. Wu, et al., Detection and seasonal variations of Huanglongbing disease in navel orange trees using direct lonization mass spectrometry, J. Agric. Food. Chem. , 67 (2019): 2265–2271. |
| [7] | C. Yang, H. Chen, H. Chen, et al., Antioxidant and anticancer activities of essential oil from Gannan navel orange peel, Molecules, 22 (2017): 1931–1400. |
| [8] | G. Rao, L. Huang, M. H. Liu, et al., Identification of Huanglongbing-infected navel oranges based on laser-induced breakdown spectroscopy combined with different Chemometric methods, Appl. Optics., 57 (2018): 8738–8742. |
| [9] | Y. H. Liu and J. H. Tsai, Effects of temperature on biology and life table parameters of the Asian citrus psyllid, Diaphorina Citri Kuwayama (Homoptera: Psyllidae), Ann. Appl. Biol., 137 (2000): 201–206. |
| [10] | T. H. Hung, S. C. Hung, C. N. Chen, et al., Detection by PCR of Candidatus Liberibacter asiaticus,the bacterium causing citrus huanglongbing in vector psyllids: application to the study of vector pathogen relationships, Plant Pathol., 53 (2004): 96–102. |
| [11] | F. Wu, Y. J. Cen, X. L. Deng, et al., Movement of Diaphorina citri(Hemiptera: Liviidae) adults between huanglongbing infected and healthy citrus, Flor. Entomol., 98 (2015): 410–416. |
| [12] | W. Shen, S. E. Halbert, E. Dickstein, et al., Occurrence and ingrove distribution of citrus huang- longbing in north central Florida, J. Plant. Pathol., 95 (2003): 361–371. |
| [13] | K. L. Manjunath, S. E. Halbert, C. Ramadugu, et al., Detection of Candidatus Liberibacter asiaticus in Diaphorina citri and its importance in the management of citrus huanglongbing in Florida, Phytopathology, 98 (2008): 387–396. |
| [14] | E. E. G. Cardwell, L. L. Stelinski and P. A. Stansly, Biology and man agement of Asian Citrus Psyllid,vector of the Huanglongbing pathogens, Annu. Rev. Entomol., 58 (2013): 413–432. |
| [15] | X. Z. Meng and Z. Q. Li, The dynamics of plant disease models with continuous and impulsive cultural control strategies, J. Theor. Biol., 266 (2010): 29–40. |
| [16] | X. S. Zhang and J. Holt, Mathematical models of cross protection in the epidemiology of plant- virus disease, Anal. Theo. Plant. Pathol., 91 (2001): 924–934. |
| [17] | S. J. Gao, L. J. Xia, Y. Liu, et al., A plant virus disease model with periodic environment and pulse roguing, Stud. Appl. Math., 136 (2016): 357–381. |
| [18] | S. Y. Tang, Y. N. Xiao and R. A. Cheke, Dynamical analysis of plant disease models with cultural control strategies and economic thresholds, Math. Comput. Simul., 80 (2010): 894–921. |
| [19] | M. S. Chan and M. J. Jeger, An analytical model of plant virus disease dynamics with roguing and replanting, J. Appl. Ecol., 31 (1994): 413–427. |
| [20] | C. Chiyaka, B. H. Singer, S. E. Halbert, et al., Modeling huanglongbing transmission within a citrus tree, PNAS, 109 (2012): 12213–12218. |
| [21] | R. A. Taylor, E. A. Mordecai, C. A. Gilligan, et al., Mathematical models are a powerful method to understand and control the spread of Huanglongbing, Peer J, 4 (2016): DOI 10.7717/peerj.2642. |
| [22] | K. Jacobsen, J. Stupiansky and S. S. Pilyugin, Mathematical modeling of citrus groves infected by Huanglongbing, Math. Biosci. Eng, 10 (2013): 705–728. |
| [23] | R. G. dA. Vilamiu, S. Ternes, G. A. Braga, et al., A model for Huanglongbing spread between citrus plants including delay times and human intervention, AIP. Conf. Proc., 1479 (2012): 2315–2319. |
| [24] | S. J. Gao, L. Luo, S. X. Yan, et al., Dynamical behavior of a novel impulsive switching model for HLB with seasonal fluctuations, Complexity, 2018 (2018): 1–11. |
| [25] | J.A. Lee, S. E. Halbert, W. O. Dawson, et al., Asymptomatic spread of Huanglongbing and implications for disease control, Proc. Natl. Acad. Sci. USA., 112 (2015): 7605–7610. |
| [26] | M. Parry, G. J. Gibson, S. Parnell, et al., Bayesian inference for an emerging arboreal epidemic in the presence of control, Proc. Natl. Acad. Sci. USA., 111 (2014): 6258–6262. |
| [27] | S. E. Halbert and K. L. Manjunath, Asian citrus psyllids (Sternorrhyncha: Psyllidae) and greening disease of citrus: A literature review and assessment of risk in Florida, Flor. Entomol., 87 (2004): 330–353. |
| [28] | W. D. Wang and X. Q. Zhao, Threshold dynamics for compartmental epidemic models in periodic environments, J. Dyn. Differ. Equ., 20 (2008): 699–717. |
| [29] | P. V. D. Driessche and J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002): 29–48. |
| [30] | H. R. Thieme, Convergence results and poincare-bendixson trichotomy for asymptotically au-tonomous differential equations, J. Math. Biol., 30 (1992): 755–763. |
| [31] | P. V. D. Driessche and J. Watmough, Reproductive numbers and subthreshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002): 29–48. |
| [32] | D. Xu and X. Q. Zhao, Dynamics in a periodic competitive model with stage structure, J. Math. Anal. Appl., 311 (2005): 417–438. |
| [33] | X. Zhao, Dynamical Systems in Population Biology, Springer, New York, NY, USA, 2003. |
| [34] | T. K. Kar and A. Batabyal, Stability analysis and optimal control of an SIR epidemic model with vaccination, Biosystems, 104 (2011): 127–135. |
| [35] | L. Y. Pang, S. G. Ruan, S. H. Liu, et al. , Transmission dynamics and optimal control of measles epidemics, Appl. Math. Comput., 256 (2015): 131–147. |
| [36] | M. A. Khan, R. Khan, Y. Khan, et al., A mathematical analysis of Pine Wilt disease with variable population size and optimal control strategies, Chaos, Solitons and Fract., 108 (2018): 205–217. |
| [37] | W. H. Fleming and R. W. Rishel, Deterministic and Stochastic Optimal Control, Springer-Verlag, New York, Berlin, 1975. |
| [38] | D. L. Lukes, Differential equations: Classical to Controlled, Mathematics in Science and Engi-neering, Academic Press, New York, 1982. |
| [39] | S. Lenhart and J. T. Workman, Optimal control applied to biological models, Mathematical and Computational Biology Series, Chapman & Hall/CRC Press, London/Boca Raton, 2007. |
| [40] | X. Zhang, Y. Zhao and A. Neumann, Partial immunity and vaccination for influenza, J. Comput. Biol., 17 (2009): 1689–1696. |
| [41] | T. Gottwald, Current epidemiological understanding of citrus Huanglongbing, Annu. Rev. Phy-topathol., 48 (2010): 119–139. |
| [42] | K. S. P. Stelinski, R. H. Brlansky, T. A. Ebert, et al., Transmission parameters for Candidatus Liberibacter asiaticus by Asian citrus psyllid (Hemiptera: Psyllidae), J. Econ. Entomol., 103 (2010): 1531–1541. |
| [43] | M. E. Rogers, General pest management considerations, Citrus. Ind., 89 (2008): 12–15. |
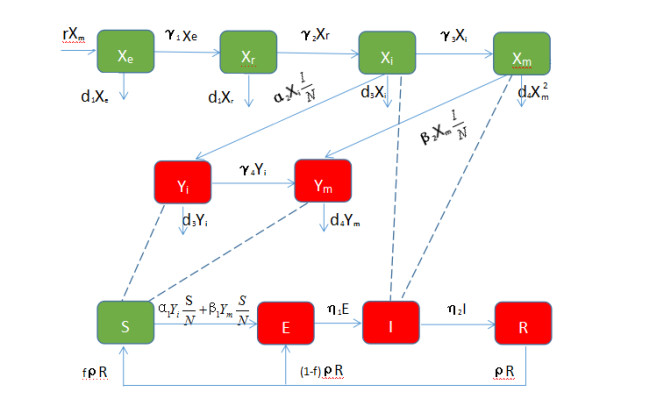
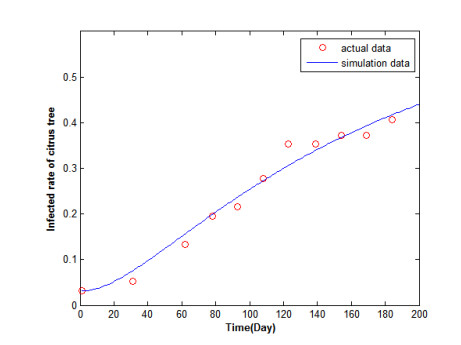
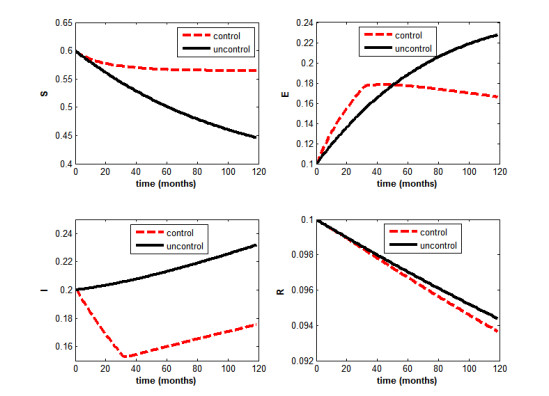
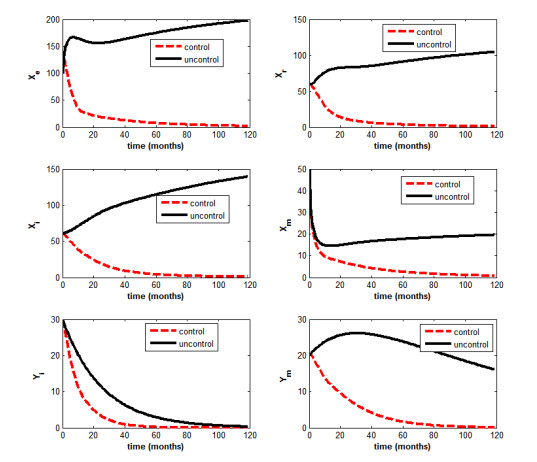
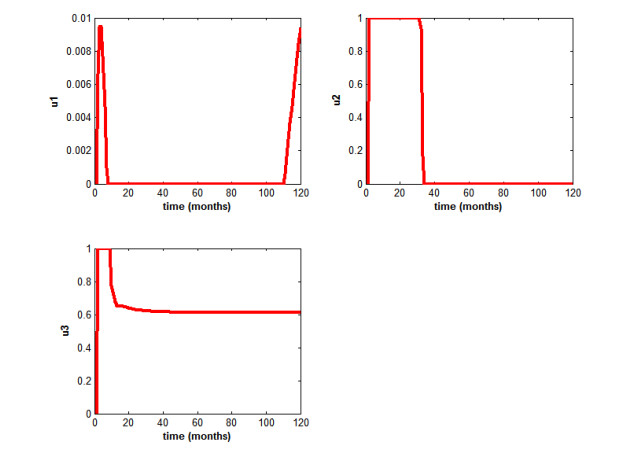
Figures(5) / Tables(2)

Yunbo Tu, Shujing Gao, Yujiang Liu, Di Chen, Yan Xu. Transmission dynamics and optimal control of stage-structured HLB model[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 5180-5205. doi: 10.3934/mbe.2019259







 DownLoad:
DownLoad: