Citation: Jiazhe Lin, Rui Xu, Xiaohong Tian. Transmission dynamics of cholera with hyperinfectious and hypoinfectious vibrios: mathematical modelling and control strategies[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 4339-4358. doi: 10.3934/mbe.2019216

| [1] | Cholera Fact Sheets, World Health Organization, August 2017. Available from: https://www.who.int/en/news-room/fact-sheets/detail/cholera. |
| [2] | J. H. Tien and D. J. D. Earn, Multiple transmission pathways and disease dynamics in a waterborne pathogen model, Bull. Math. Biol., 72 (2010), 1506–1533. |
| [3] | X. Zhou and J. Cui, Threshold dynamics for a cholera epidemic model with periodic transmission rate, Appl. Math. Model., 37 (2013), 3093–3101. |
| [4] | D. Posny, J. Wang, Z. Mukandavire, et al., Analyzing transmission dynamics of cholera with public health interventions, Math. Biosci., 264 (2015), 38–53. |
| [5] | Y. Wang and J. Cao, Global stability of general cholera models with nonlinear incidence and removal rates, J. Frankl. Inst., 352 (2015), 2464–2485. |
| [6] | J. P. Tian and J. Wang, Global stability for cholera epidemic models, Math. Biosci., 232 (2011), 31–41. |
| [7] | D. M. Hartley, J. G. Morris and D. L. Smith, Hyperinfectivity: A critical element in the ability of V. cholerae to cause epidemics? PLoS Med., 3 (2006), 63–69. |
| [8] | R. L. M. Neilan, E. Schaefer, H. Gaff, et al., Modeling optimal intervention strategies for cholera, Bull. Math. Biol., 72 (2010), 2004–2018. |
| [9] | Z. Mukandavire, S. Liao, J. Wang, et al., Estimating the reproductive numbers for the 2008–2009 cholera outbreaks in Zimbabwe, PNAS, 108 (2011), 8767–8772. |
| [10] | T. K. Sengupta, R. K. Nandy, S. Mukhopadhyay, et al., Characterization of a 20-kDa pilus protein expressed by a diarrheogenic strain of non-O1/non-O139 vibrio cholerae, FEMS Microbiol. Lett., 160 (1998), 183–189. |
| [11] | C. T. Codec ¸o, Endemic and epidemic dynamics of cholera: the role of the aquatic reservoir, BMC Infect. Dis., 1 (2001), 1–14. |
| [12] | C. Modnak, A model of cholera transmission with hyperinfectivity and its optimal vaccination control, Int. J. Biomath., 10 (2017), 1750084. |
| [13] | A.P.Lemos-Paião, C.J.SilvaandD.F.M.Torres, A cholera mathematical model with vaccination and the biggest outbreak of world's history, AIMS Mathematics, 3 (2018), 448–463. |
| [14] | J. K. Hale and S. Verduyn Lunel, Introduction to Functional Differential Equations, 1st edition, Springer-Verlag, New York, 1993. |
| [15] | P. van den Driessche and J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002), 29–48. |
| [16] | O. Sharomi and T. Malik, Optimal control in epidemiology, Ann. Oper. Res., 251 (2017), 55–71. |
| [17] | J. K. K. Asamoah, F. T. Oduro, E. Bonyah, et al., Modelling of rabies transmission dynamics using optimal control analysis, J. Appl. Math., 12 (2017), 1–23. |
| [18] | L. Cesari, Optimization-Theory and Applications. Problems with ordinary differential equations, in: Applications of Mathematics, vol. 17, Springer-Verlag, New York, 1983. |
| [19] | W. H. Fleming and R. W. Rishel, Deterministic and Stochastic Optimal Control, 1st edition, Springer-Verlag, New York, 1975. |
| [20] | L. Pontryagin, V. Boltyanskii, R. Gramkrelidze, et al., The Mathematical Theory of Optimal Processes, John Wiley & Sons, 1962. |
| [21] | C. J. Silva and D. F. M. Torres, Optimal control for a tuberculosis model with reinfection and post-exposure interventions, Math. Biosci., 244 (2013), 154–164. |
| [22] | M. McAsey, L. Mou and W. Han, Convergence of the forward-backward sweep method in optimal control, Comput. Optim. Appl., 53 (2012), 207–226. |
| [23] | O. Zakary, A. Larrache, M. Rachik, et al., Effect of awareness programs and travel-blocking operations in the control of HIV/AIDS outbreaks: a multi-domains SIR model, Adv. Differ. Equ., 2016 (2016), 169. |
| [24] | C. Modnak, J. Wang and Z. Mukandavire, Simulating optimal vaccination times during cholera outbreaks, Int. J. Biomath., 7 (2014), 1450014. |
| [25] | J. Wang and C. Modnak, Modeling cholera dynamics with controls, Canadian Appl. Math. Quart., 19 (2011), 255–273. |
| [26] | D.S.Merrell, S.M.Butler, F.Qadri, et al., Host-induced epidemic spread of the cholera bacterium, Nature, 417 (2002), 642–645. |
| [27] | S. Lenhart and J. T. Workman, Optimal control applied to biological models, Chapman & Hall/CRC Press, Boca Raton, 2007. |
| [28] | Global Task Force on Cholera Control, Cholera Country Profile: Haiti, World Health Organization, May 2011. Available from: https://www.who.int/cholera/countries/HaitiCountryProfileMay2011.pdf. |
| [29] | A. P. Lemos-Paião, C. J. Silva and D. F.M. Torres, An epidemic model for cholera with optimal control treatment, J. Comput. Appl. Math., 318 (2017), 168–180. |
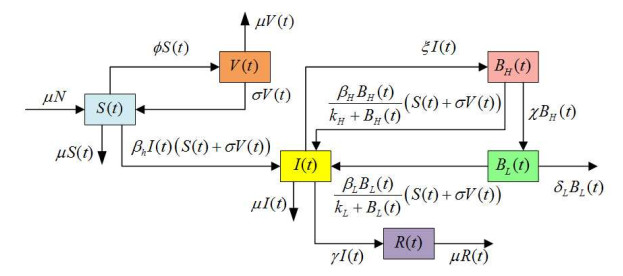
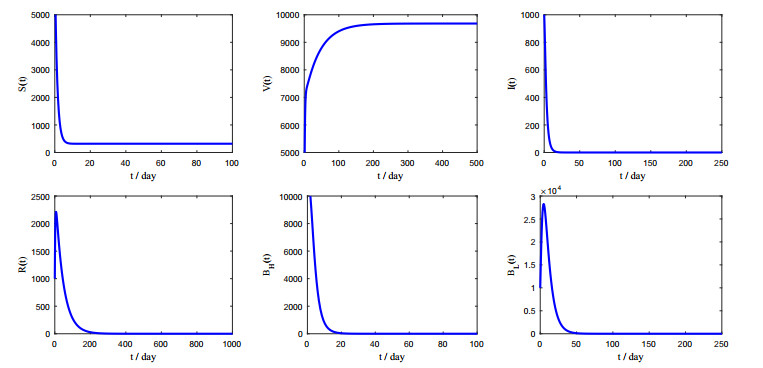
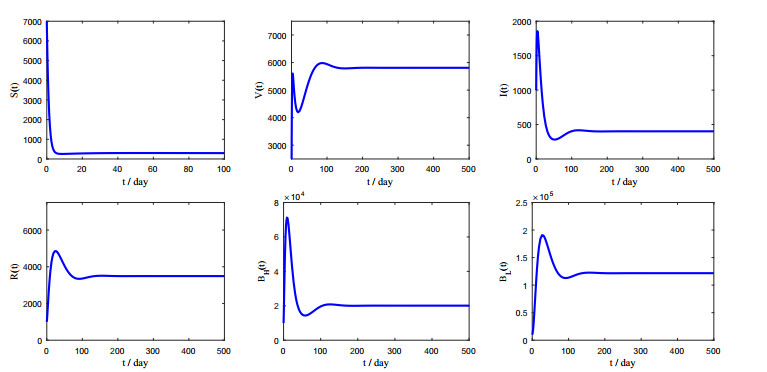
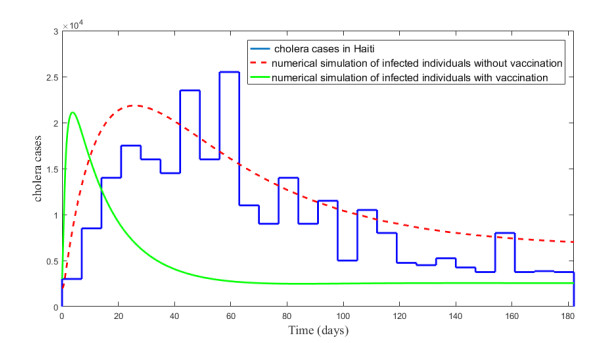
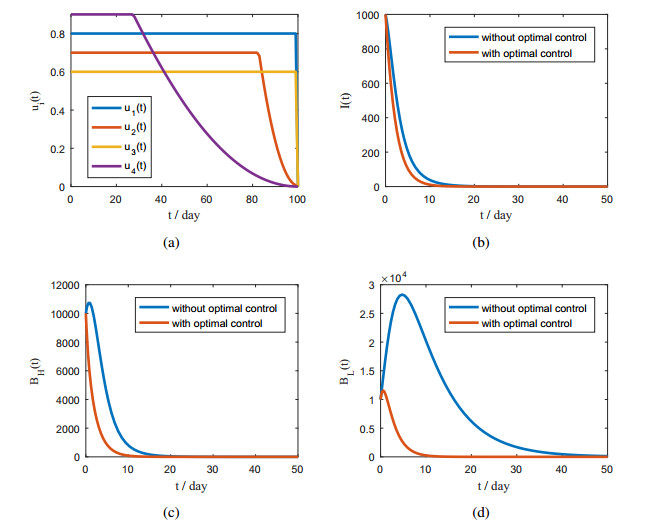
Figures(5) / Tables(2)

Jiazhe Lin, Rui Xu, Xiaohong Tian. Transmission dynamics of cholera with hyperinfectious and hypoinfectious vibrios: mathematical modelling and control strategies[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 4339-4358. doi: 10.3934/mbe.2019216







 DownLoad:
DownLoad: