Citation: Ning Bai, Juan Zhang , Li Li, Zhen Jin. Evaluating the effect of virus mutation on the transmission of avianinfluenza H7N9 virus in China based on dynamical model[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 3393-3410. doi: 10.3934/mbe.2019170

| [1] | R. Gao, B. Cao, Y. Hu, et al, Human infection with a novel avian-origin influenza A (H7N9) virus, N. Engl. J. Med., 368 (2013), 1888–1897. |
| [2] | Q. Li, L. Zhou, M. Zhou, et al, Epidemiology of Human Infections with Avian Influenza A (H7N9) Virus in China, N. Engl. J. Med., 370 (2014), 520–532. |
| [3] | L. Fang, X. Li, K. Liu, et al, Mapping Spread and Risk of Avian Influenza A (H7N9) in China, Sci. Rep., 3 (2013), 2722. |
| [4] | Avian Influenza Report, Report of the government of the Hong Kong Special Administrative Region, Available from: https://www.chp.gov.hk/sc/resources/29/332.html. |
| [5] | X. Wang, H. Jiang, P. Wu, et al, Epidemiology of avian influenza A H7N9 virus in human beings across five epidemics in mainland China, 2013-17: an epidemiological study of laboratory- confirmed case series, Lancet Infect. Dis., 17 (2017), 822–832. |
| [6] | A. Badulak, Human Infections with the Emerging Avian Influenza A H7N9 Virus from Wet Market Poultry: Clinical Analysis and Characterization of Viral Genome, J. Emerg. Med., 45 (2013), 484. |
| [7] | Z. Chen, H. Liu, J. Lu, et al, Asymptomatic, Mild, and Severe Influenza A(H7N9) Virus Infection in Humans, Guangzhou, China, Emerg. Infect. Dis., 20 (2014), 1535–1540. |
| [8] | Human infection with avian influenza A(H7N9) virus - China, Report of World Health Organiza- tion, Available from: http://www.who.int/csr/don/27-february-2017-ah7n9-china/ en/. |
| [9] | W. Zhu, J. Zhou, Z. Li, et al, Biological characterisation of the emerged highly pathogenic avian influenza (HPAI) A(H7N9) viruses in humans, in mainland China, 2016 to 2017, Euro. Surveill., 22 (2017). |
| [10] | L. Yang , W. Zhu, X. Li, et al, Genesis and Spread of Newly Emerged Highly Pathogenic H7N9 Avian Viruses in Mainland China, J. Virol., 91(2017), JVI.01277-17. |
| [11] | C. Quan, W. Shi, Y. Yang, et al, New threats of H7N9 influenza virus: the spread and evolution of highly and low pathogenic variants with high genomic diversity in Wave Five, J. Virol., 92 (2018), JVI.00301-18. |
| [12] | Official Veterinary Bulletin, Report of Ministry of Agriculture of the People's Republic of China, Available from: http://jiuban.moa.gov.cn/zwllm/tzgg/gb/sygb/. |
| [13] | R. Mu and Y. Yang, Global Dynamics of an Avian Influenza A(H7N9) Epidemic Model with Latent Period and Nonlinear Recovery Rate, Comput. Math. Methods Med., 2018 (2018), 1–11. |
| [14] | Y. Chen and Y. Wen, Global dynamic analysis of a H7N9 avian-human influenza model in an outbreak region, J. Theor. Biol., 367 (2015), 180–188. |
| [15] | J. Zhang, Z. Jin, G. Sun, et al, Determination of original infection source of H7N9 avian influenza by dynamical model, Sci. Rep., 4 (2014), 4846. |
| [16] | Z. Liu and C. Fang, A modeling study of human infections with avian influenza A H7N9 virus in mainland China, Int. J. Infect. Dis., 41 (2015), 73–78. |
| [17] | Y. Xing, L. Song, G. Sun, et al, Assessing reappearance factors of H7N9 avian influenza in China, Appl. Math. Comput., 309 (2017), 192–204. |
| [18] | M. Kang, L. Ehy, W. Guan, et al, Epidemiology of human infections with highly pathogenic avian influenza A(H7N9) virus in Guangdong, 2016 to 2017, Euro. Surveill., 22 (2017). |
| [19] | W. Su, K. Cheng, D. Chu, et al, Genetic analysis of H7N9 highly pathogenic avian influenza virus in Guangdong, China, 2016C2017, J. Infect., 76 (2018). |
| [20] | L. Cao, K.B. Li, Y. Liu, et al, Surveillance for circulation and genetic evolution of avian influenza A (H7N9) virus in poultry markets in Guangzhou, 2014 - 2017, Dis. Survell., 33 (2018), 897–901. |
| [21] | C. Li, R.Ren, D. Li, et al, Epidemic and death case analysis on human infection with highly pathogenic avian influenza A (H7N9) virus in the mainland of China, 2016 - 2018, Dis. Survell., 33 (2018), 985–989. |
| [22] | D. Wu, S. Zou, T. Bai, et al, Poultry farms as a source of avian influenza A (H7N9) virus reassortment and human infection, Sci. Rep., 5 (2015), 7630. |
| [23] | K.Tan, S. Jacob, K. Chan, et al, An overview of the characteristics of the novel avian influenza A H7N9 virus in humans, Front Microbiol., 6 (2015), 140. |
| [24] | Y. Bi, J. Liu, H. Xiong, et al, A new reassortment of influenza A (H7N9) virus causing human infection in Beijing, 2014, Sci. Rep., 6 (2016), 26624. |
| [25] | A. Handel, J. Brown, D. Stallknecht, et al, A Multi-scale Analysis of Influenza A Virus Fit- ness Trade-offs due to Temperature-dependent Virus Persistence, PLoS Comput. Biol., 9 (2013), e1002989. |
| [26] | Guangdong statistical yearbook, Report of Guangdong statistical information network, Available from: http://www.gdstats.gov.cn/tjsj/gdtjnj/. |
| [27] | Nation data, Report of Nation Bureau of Statistics of China, Available from: http://data. stats.gov.cn/easyquery.htm?cn=E0103. |
| [28] | J. Ma and Z. Ma, Epidemic threshold conditions for seasonally forced SEIR models, Math. Biosci. Eng., 3 (2017), 161–172. |
| [29] | X. Hu, Threshold dynamics for a Tuberculosis model with seasonality, Math. Biosci. Eng., 9 (2011), 111–122. |
| [30] | W. Wang and X. Zhao, Threshold Dynamics for Compartmental Epidemic Models in Periodic Environments, J. Dyn. Differ. Equ., 20 (2008), 699–717. |
| [31] | P. van den Driessche and J. Watmough, Reproduction numbers and sub-threshold endemic equi- libria for compartmental models of disease transmission, Math. Biosci., 180 (2002), 29–48. |
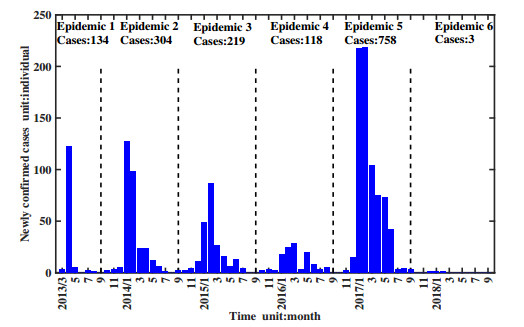
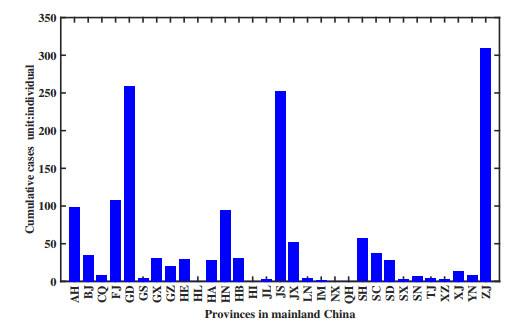
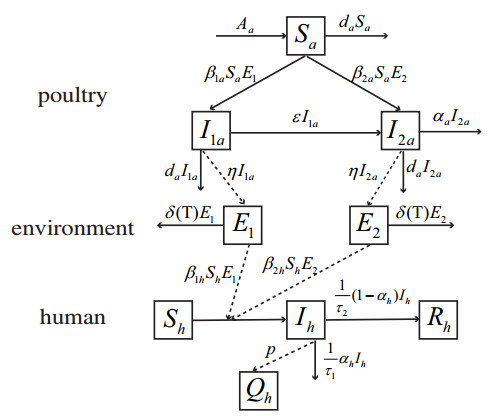
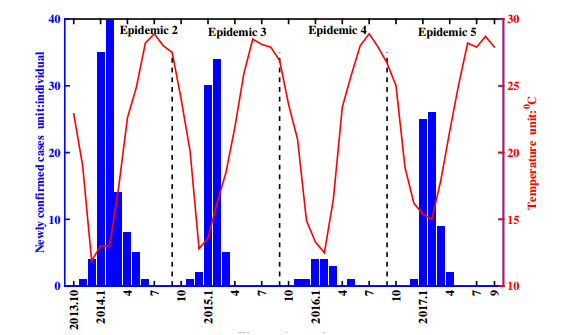
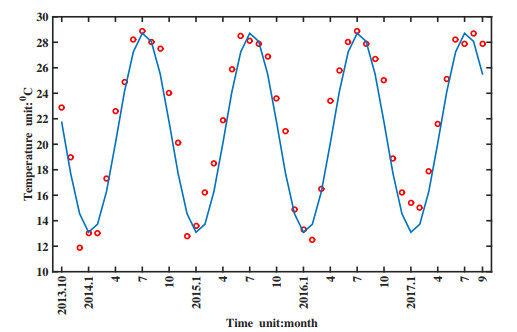
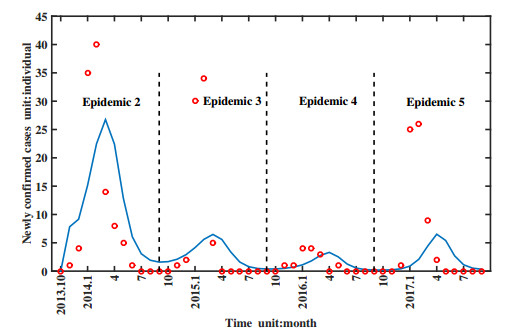
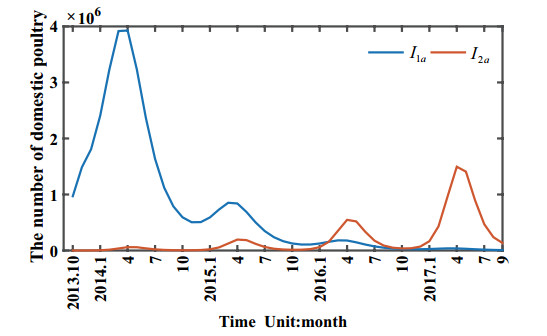
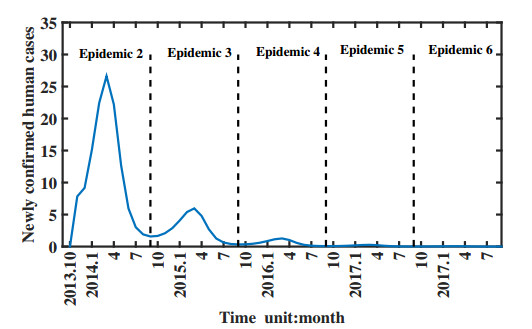
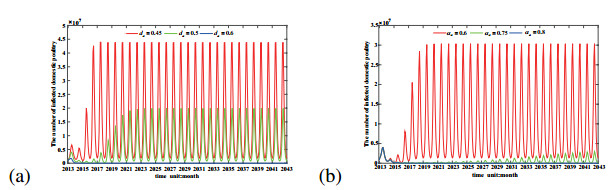
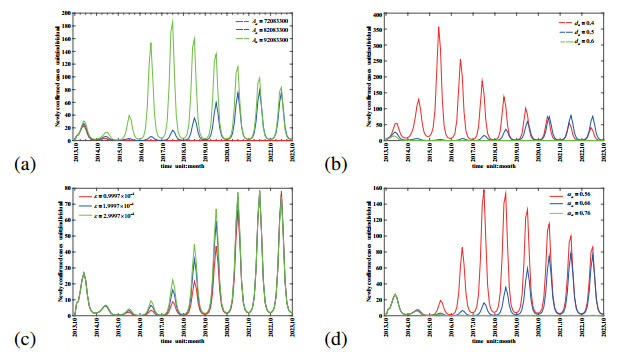
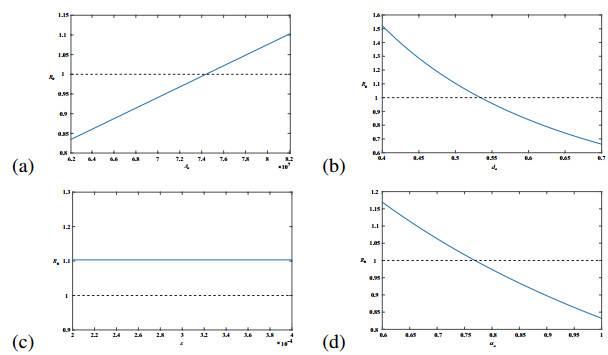
Figures(11) / Tables(1)

Ning Bai, Juan Zhang , Li Li, Zhen Jin. Evaluating the effect of virus mutation on the transmission of avian influenza H7N9 virus in China based on dynamical model[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 3393-3410. doi: 10.3934/mbe.2019170







 DownLoad:
DownLoad: