Citation: Gordon Akudibillah, Abhishek Pandey, Jan Medlock. Optimal control for HIV treatment[J]. Mathematical Biosciences and Engineering, 2019, 16(1): 373-396. doi: 10.3934/mbe.2019018

| [1] | F. B. Agusto, N. Marcus, and K. O. Okosun, Application of optimal control to the epidemiology of malaria. Electron. J. Differ. Eq., 81 (2012), 1–22. |
| [2] | G. Akudibillah, A. Pandey and J. Medlock, Maximizing the benefits of art and prep in resourcelimited settings. Epidemiol. Infect., 145 (2017), 942–956. |
| [3] | M. Alary, L. Mukenge-Tshibaka, F. Bernier, N. Geraldo, C. M. Lowndes, H. Meda, C. A. B. Gnintoungb`e, S. Anagonou and J. R. Joly. Decline in the prevalence of HIV and sexually transmitted diseases among female sex workers in Cotonou, Benin, 1993–1999. AIDS, 16 (2002), 463–470. |
| [4] | R. M. Andersen and R. M. May, Epidemiological parameters of HIV transmission. Nature, 333 (1988), 514–519. |
| [5] | R. E. Berger. Re: Effectiveness and safety of tenofovir gel, an antiretroviral microbicide, for the prevention of HIV infection in women. J. Urol., 185 (2011), 1729. |
| [6] | M. H. A. Biswas, L. T. Paiva, and M. D. R. De Pinho. A SEIR model for control of infectious diseases with constraints. Math. Biosci. Eng., 11 (2013), 761–784. |
| [7] | N. Bráu, M. Salvatore, C. F. Ríos-Bedoya, A. Fernández-Carbia, F. Paronetto, J. F. Rodríguez- Orengo, and M. Rodríguez-Torres. Slower fibrosis progression in HIV/HCV-coinfected patients with successful HIV suppression using antiretroviral therapy. J. Hepatol., 44 (2006), 47–55. |
| [8] | S. Butler, D. Kirschner, and S. Lenhart. Optimal control of chemotherapy affecting the infectivity of hiv. Ann Arbor, 1001 (1997), 48109–0620. |
| [9] | T. Clayton, S. Duke-Sylvester, L. J. Gross, S. Lenhart, and L. A. Real. Optimal control of a rabies epidemic model with a birth pulse. J. Biol. Dyn., 4 (2010), 43–58. |
| [10] | S. M. Cleary, D. McIntyre, and A. M. Boulle. The cost-effectiveness of antiretroviral treatment in Khayelitsha, South Africa–a primary data analysis. Cost Effectiveness and Resource Allocation, 4 (2006), 20. |
| [11] | M. S. Cohen, Y. Q. Chen, M. McCauley, T. Gamble, M. C. Hosseinipour, N. Kumarasamy, J. G. Hakim, J. Kumwenda, B. Grinsztejn, J. H. Pilotto, et al. Prevention of HIV-1 infection with early antiretroviral therapy. New Engl. J. Med., 365 (2011), 493–505. |
| [12] | M. D. R. De Pinho, I. Kornienko, and H. Maurer. Optimal control of a SEIR model with mixed constraints and L1 cost. In A. P. Moreira, A. Matos, and G. Veiga, eds., CONTROLO'2014 – Proceedings of the 11th Portuguese Conference on Automatic Control, pp. 135–145. Springer, New York, 2015. ISBN 978-3-319-10379-2. doi:10.1007/978-3-319-10380-8 14. |
| [13] | A. K. Dixit and R. S. Pindyck. Investment Under Uncertainty. Princeton University Press, Princeton, New Jersey, 1994. ISBN 978-0691034102. |
| [14] | W. Fleming and R. Rishel. Deterministic and Stochastic Optimal Control. Springer New York, 1975. ISBN 978-1-4612-6382-1. |
| [15] | M. Gold, J. Siegel, L. Russell, and M. Weinstein. Cost-Effectiveness in Health and Medicine. Oxford University Press, New York, 1996. ISBN 978-0195108248. |
| [16] | K. Hattaf and N. Yousfi. Two optimal treatments of HIV infection model. World Journal of Modelling and Simulation, 8 (2012), 27–35. |
| [17] | L. F. Johnson, T. M. Rehle, S. Jooste, and L.-G. Bekker. Rates of HIV testing and diagnosis in South Africa: successes and challenges. AIDS, 29 (2015), 1401–1409. |
| [18] | H. R. Joshi, Optimal control of an hiv immunology model, Optim. contr. appl. met., 23 (2002), 199–213. |
| [19] | E. Jung, S. Iwami, Y. Takeuchi, and T.-C. Jo, Optimal control strategy for prevention of avian influenza pandemic, J. Theor. Biol., 260 (2009), 220–229. |
| [20] | D. Kirschner, S. Lenhart, and S. Serbin, Optimal control of the chemotherapy of HIV, J. Math. Biol., 35 (1997), 775–792. |
| [21] | S. Lenhart, E. Jung, and Z. Feng, Optimal control of treatments in a two-strain tuberculosis model, Discrete Cont. Dyn.-B, 2 (2002), 473–482. |
| [22] | S. Lenhart, V. Protopopescu, E. Jung, and C. Babbs, Optimal control for a standard CPR model. Nonlinear Anal. Theor., 63 (2005), e1391–e1397. |
| [23] | J. A. Levy, Mysteries of HIV: challenges for therapy and prevention, Nature, 333 (1988), 519–522. |
| [24] | I. M. Longini, W. S. Clark, R. H. Byers, J. W. Ward, W. W. Darrow, G. F. Lemp, and H. W. Hethcote, Statistical analysis of the stages of HIV infection using a Markov model, Stat. Med., 8 (1989), 831–843. |
| [25] | G. Marks, N. Crepaz, J.W. Senterfitt, and R. S. Janssen, Meta-analysis of high-risk sexual behavior in persons aware and unaware they are infected with hiv in the united states: implications for hiv prevention programs, J. Acquir. Immune Defic. Syndr., 39 (2005), 446–453. |
| [26] | Médecins Sans Fronti`eres. Untangling the web of antiretroviral price reductions: 14th edition July 2011. 2011. Available from: http://d2pd3b5abq75bb.cloudfront.net/2012/07/16/14/ 42/23/52/UTW_14_ENG_July2011.pdf. |
| [27] | R. M. Neilan and S. Lenhart, An introduction to optimal control with an application in disease modeling. In A. B. Gumel and S. Lenhart, eds., Modeling Paradigms and Analysis of Disease Transmission Models, vol. 75 of DIMACS Series in Discrete Mathematics, pp. 67–81. American Mathematical Society, Providence, Rhode Island, 2010. ISBN 978-0-8218-4384-0. Available from: https://www.researchgate.net/profile/Rachael_Miller_ Neilan/publication/265363622_An_Introduction_to_Optimal_Control_with_an_ Application_in_Disease_Modeling/links/5597d5e908ae21086d22b532.pdf. |
| [28] | K. Okosun, O. Makinde, and I. Takaidza, Impact of optimal control on the treatment of HIV/AIDS and screening of unaware infectives, Appl. Math. Model., 37 (2013), 3802–3820. |
| [29] | L. S. Pontryagin. Mathematical Theory of Optimal Processes. CRC Press, Boca Raton, Florida, 1987. ISBN 9782881240775. |
| [30] | L. Simbayi, O. Shisana, T. Rehle, D. Onoya, S. Jooste, N. Zungu, and K. Zuma. South African National HIV Prevalence, Incidence and Behaviour Survey, 2012. HSRC Press, Cape Town, South Africa, 2014. ISBN 978-0-7969-2483-4. Available from: http://www.hsrc.ac.za/ en/research-data/ktree-doc/15031. |
| [31] | Statistics South Africa. Mid-year population estimates 2014, 2014. Available from: http:// www.statssa.gov.za/publications/P0302/P03022014.pdf. |
| [32] | UNAIDS. Global aids update. Accessed 11 Jun. 2017. Available from: http://www.who.int/ hiv/pub/arv/global-AIDS-update-2016_en.pdf. |
| [33] | UNAIDS. AIDSinfo j UNAIDS. Accessed 25 Sept 2015. Available from: http://www.unAIDS. org/en/dataanalysis/datatools/AIDSinfo. |
| [34] | A. I. van Sighem, M. A. van de Wiel, A. C. Ghani, M. Jambroes, P. Reiss, I. C. Gyssens, K. Brinkman, J. M. Lange, and F. de Wolf, Mortality and progression to AIDS after starting highly active antiretroviral therapy, AIDS, 17 (2003), 2227–2236. |
| [35] | X. Wang. Solving optimal control problems with MATLAB-indirect methods. 2009. Available from: http://www4.ncsu.edu/~xwang10/document/Solving%20optimal%20control% 20problems%20with%20MATLAB.pdf. |
| [36] | M. J.Wawer, R. H. Gray, N. K. Sewankambo, D. Serwadda, X. Li, O. Laeyendecker, N. Kiwanuka, G. Kigozi, M. Kiddugavu, T. Lutalo, F. Nalugoda, F. Wabwire-Mangen, M. P. Meehan, and T. C. Quinn, Rates of HIV-1 transmission per coital act by stage of HIV-1 infection, in Rakai, Uganda, J. Infect. Dis., 191 (2005), 1403–1409. |
| [37] | M. C. Weinstein, J. E. Siegel, M. R. Gold, M. S. Kamlet and L. B. Russell, Recommendations of the panel on cost-effectiveness in health and medicine, Jama, 276 (1996), 1253–1258. |
| [38] | D. P. Wilson, A. Hoare, D. G. Regan and M. G. Law, Importance of promoting HIV testing for preventing secondary transmissions: modelling the Australian HIV epidemic among men who have sex with men, Sexual Health, 6 (2009), 19. |
| [39] | World Health Organization, Interim WHO Clinical Staging of HIV/AIDS and HIV/AIDS Case Definitions For Surveillance: African Region, 2005. Available from: http://www.who.int/ hiv/pub/guidelines/clinicalstaging.pdf. |
| [40] | World Health Organization, March 2014 Supplement to the 2013 Consolidated Guidelines on the Use of Antiretroviral Drugs for Treating and Preventing HIV Infection: Recommendations for a Public Health Approach, 2014. Available from: http://apps.who.int/iris/bitstream/ 10665/104264/1/9789241506830_eng.pdf. |
| [41] | T. T. Yusuf and F. Benyah, Optimal strategy for controlling the spread of HIV/AIDS disease: a case study of South Africa, J. Biol. Dyn., 6 (2012), 475–494. |
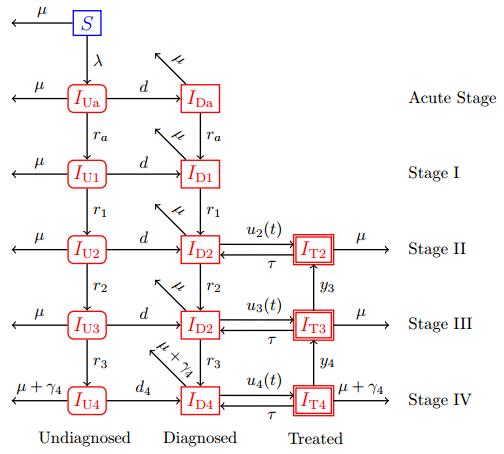
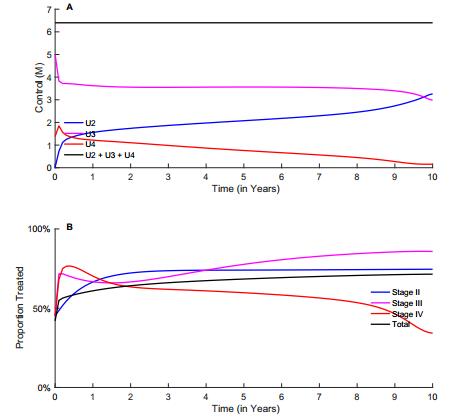
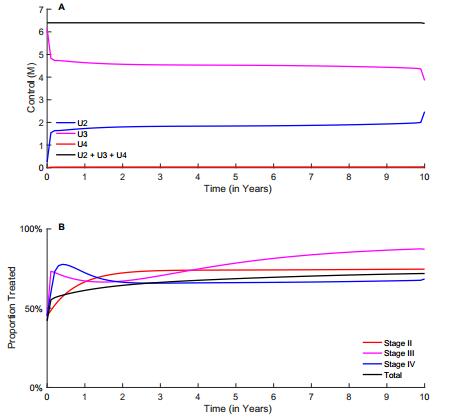
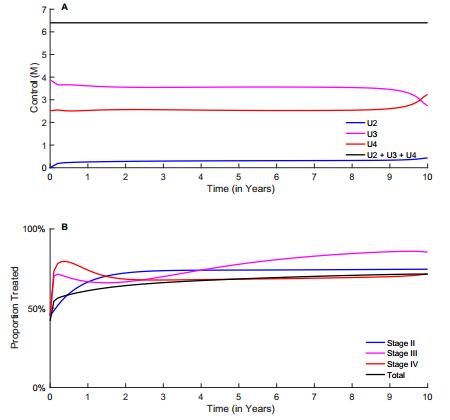
Figures(10) / Tables(2)

Gordon Akudibillah, Abhishek Pandey, Jan Medlock. Optimal control for HIV treatment[J]. Mathematical Biosciences and Engineering, 2019, 16(1): 373-396. doi: 10.3934/mbe.2019018







 DownLoad:
DownLoad: