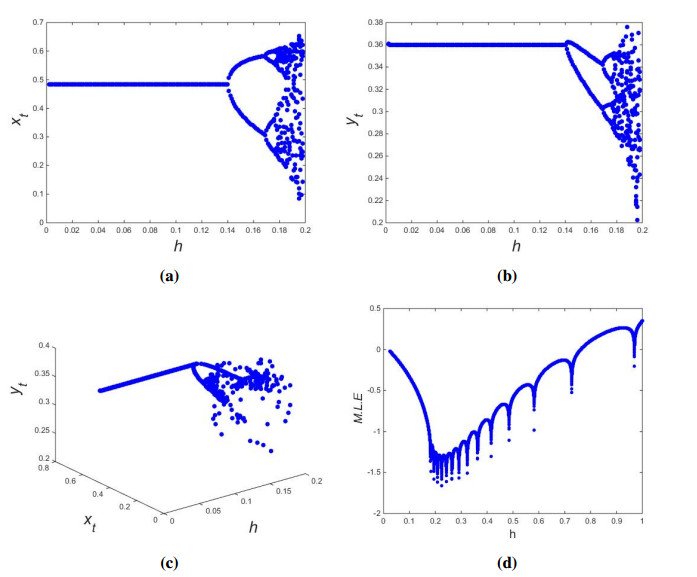
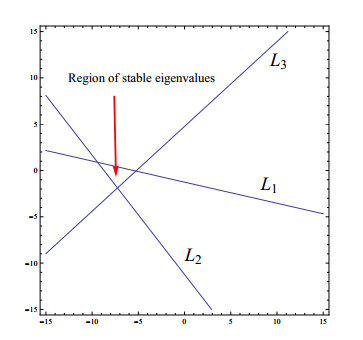
In this paper, we explore the existence of fixed points, local dynamics at fixed points, bifurcations and chaos of a discrete prey-predator fishery model with harvesting. More specifically, it is proved that, for all involved parameters, the model has trivial fixed point, but it has semitrivial and interior fixed points under definite parametric condition(s). We study the local behavior at fixed points by applying the theory of linear stability. Furthermore, it is shown that flip bifurcation does not occur at semitrivial and trivial fixed points, but that the model undergoes Neimark-Sacker bifurcation at interior fixed point. It is also proved that, at interior fixed point, the model undergoes the flip bifurcation. By using a feedback control strategy, the chaos control is also examined. Finally, to illustrate the theoretical findings, detailed numerical simulations are provided.
Citation: A. Q. Khan, Ibraheem M. Alsulami, S. K. A. Hamdani. Controlling the chaos and bifurcations of a discrete prey-predator model[J]. AIMS Mathematics, 2024, 9(1): 1783-1818. doi: 10.3934/math.2024087
In this paper, we explore the existence of fixed points, local dynamics at fixed points, bifurcations and chaos of a discrete prey-predator fishery model with harvesting. More specifically, it is proved that, for all involved parameters, the model has trivial fixed point, but it has semitrivial and interior fixed points under definite parametric condition(s). We study the local behavior at fixed points by applying the theory of linear stability. Furthermore, it is shown that flip bifurcation does not occur at semitrivial and trivial fixed points, but that the model undergoes Neimark-Sacker bifurcation at interior fixed point. It is also proved that, at interior fixed point, the model undergoes the flip bifurcation. By using a feedback control strategy, the chaos control is also examined. Finally, to illustrate the theoretical findings, detailed numerical simulations are provided.

| [1] | M. N. Huda, F. D. T. Amijaya, I. Purnamasari, The effect of harvesting activities on prey-predator fishery model with Holling type-Ⅱ in toxicant aquatic ecosystem, Aust. J. Math. Anal. Appl., 17 (2020), 13. |
| [2] |
K. Pujaru, T. K. Kar, Impacts of predator-prey interaction on managing maximum sustainable yield and resilience, Nonlinear Anal., 25 (2020), 400–416. https://doi.org/10.15388/namc.2020.25.16657 doi: 10.15388/namc.2020.25.16657

|
| [3] |
T. K. Kar, Selective harvesting in a prey-predator fishery with time delay, Math. Comput. Modell., 38 (2003), 449–458. https://doi.org/10.1016/S0895-7177(03)90099-9 doi: 10.1016/S0895-7177(03)90099-9
|
| [4] |
H. Liu, H. Yu, C. Dai, Z. Ma, Q. Wang, M. Zhao, Dynamical analysis of an aquatic amensalism model with non-selective harvesting and Allee effect, Math. Biosci. Eng., 18 (2021), 8857–8882. https://doi.org/10.3934/mbe.2021437 doi: 10.3934/mbe.2021437
|
| [5] |
A. T. Keong, H. M. Safuan, K. Jacob, Dynamical behaviours of prey-predator fishery model with harvesting affected by toxic substances, Matematika, 34 (2018), 143–151. https://doi.org/10.11113/matematika.v34.n1.1018 doi: 10.11113/matematika.v34.n1.1018
|
| [6] |
M. Chen, R. Wu, X. Wang, Non-constant steady states and Hopf bifurcation of a species interaction model, Commun. Nonlinear Sci. Numer. Simul., 116 (2023), 106846. https://doi.org/10.1016/j.cnsns.2022.106846 doi: 10.1016/j.cnsns.2022.106846
|
| [7] |
M. Chen, R. Wu, Patterns in the predator-prey system with network connection and harvesting policy, Math. Methods Appl. Sci., 46 (2023), 2433–2454. https://doi.org/10.1002/mma.8653 doi: 10.1002/mma.8653
|
| [8] |
M. Chen, R. Wu, Dynamics of a harvested predator-prey model with predator-taxis, Bull. Malays. Math. Sci. Soc., 46 (2023), 76. https://doi.org/10.1007/s40840-023-01470-w doi: 10.1007/s40840-023-01470-w
|
| [9] |
M. Chen, H. Srivastava, Existence and stability of bifurcating solution of a chemotaxis model, Proc. Amer. Math. Soc., 151 (2023), 4735–4749. https://doi.org/10.1090/proc/16536 doi: 10.1090/proc/16536
|
| [10] |
M. Chen, H. M. Srivastava, Stability of bifurcating solution of a predator-prey model, Chaos Solitons Fract., 168 (2023), 113153. https://doi.org/10.1016/j.chaos.2023.113153 doi: 10.1016/j.chaos.2023.113153
|
| [11] |
S. M. Salman, A. M. Yousef, A. A. Elsadany, Stability, bifurcation analysis and chaos control of a discrete predator-prey system with square root functional response, Chaos Solitons Fract., 93 (2016), 20–31. https://doi.org/10.1016/j.chaos.2016.09.020 doi: 10.1016/j.chaos.2016.09.020
|
| [12] |
X. Liu, D. Xiao, Complex dynamic behaviors of a discrete-time predator-prey system, Chaos Solitons Fract., 32(2007), 80–94. https://doi.org/10.1016/j.chaos.2005.10.081 doi: 10.1016/j.chaos.2005.10.081
|
| [13] |
M. F. Elettreby, T. Nabil, A. Khawagi, Stability and bifurcation analysis of a discrete predator-prey model with mixed Holling interaction, Comput. Model. Eng. Sci., 122 (2020), 907–922. https://doi.org/10.32604/cmes.2020.08664 doi: 10.32604/cmes.2020.08664
|
| [14] |
P. K. Santra, G. S. Mahapatra, G. R. Phaijoo, Bifurcation and chaos of a discrete predator-prey model with Crowley-Martin functional response incorporating proportional prey refuge, Math. Probl. Eng., 2020 (2020), 5309814. https://doi.org/10.1155/2020/5309814 doi: 10.1155/2020/5309814
|
| [15] |
G. Zhang, L. Zhu, B. Chen, Hopf bifurcation in a delayed differential-algebraic biological economic system, Nonlinear Anal., 12 (2011), 1708–1719. https://doi.org/10.1016/j.nonrwa.2010.11.003 doi: 10.1016/j.nonrwa.2010.11.003
|
| [16] |
L. Zhang, L. Zou, Bifurcations and control in a discrete predator-prey model with strong Allee effect, Int. J. Bifurc. Chaos, 28 (2018), 1850062. https://doi.org/10.1142/S0218127418500621 doi: 10.1142/S0218127418500621
|
| [17] |
P. Chakraborty, U. Ghosh, S. Sarkar, Stability and bifurcation analysis of a discrete prey-predator model with square-root functional response and optimal harvesting, J. Biol. Syst., 28 (2020), 91–110. https://doi.org/10.1142/S0218339020500047 doi: 10.1142/S0218339020500047
|
| [18] |
D. Mua, C. Xub, Z. Liua, Y. Panga, Further insight into bifurcation and hybrid control tactics of a chlorine dioxide-iodine-malonic acid chemical reaction model incorporating delays, MATCH Commun. Math. Comput. Chem., 89 (2023), 529–566. https://doi.org/10.46793/match.89-3.529M doi: 10.46793/match.89-3.529M
|
| [19] |
C. Xu, X. Cui, P. Li, J. Yan, L. Yao, Exploration on dynamics in a discrete predator-prey competitive model involving feedback controls, J. Biol. Dyn., 17 (2023), 2220349. https://doi.org/10.1080/17513758.2023.2220349 doi: 10.1080/17513758.2023.2220349
|
| [20] |
C. Xu, Z. Liu, P. Li, J. Yan, L. Yao, Bifurcation mechanism for fractional-order three-triangle multi-delayed neural networks, Neural Process. Lett., 55 (2023), 6125–6151. https://doi.org/10.1007/s11063-022-11130-y doi: 10.1007/s11063-022-11130-y
|
| [21] |
C. Xu, D. Mu, Y. Pan, C. Aouiti, L. Yao, Exploring bifurcation in a fractional-order predator-prey system with mixed delays, J. Appl. Anal. Comput., 13 (2023), 1119–1136. https://doi.org/10.11948/20210313 doi: 10.11948/20210313
|
| [22] |
P. Li, Y. Lu, C. Xu, J. Ren, Insight into hopf bifurcation and control methods in fractional order bam neural networks incorporating symmetric structure and delay, Cogn. Comput., 15 (2023), 1825–1867. https://doi.org/10.1007/s12559-023-10155-2 doi: 10.1007/s12559-023-10155-2
|
| [23] |
C. Xu, Q. Cui, Z. Liu, Y. Pan, X. Cui, W. Ou, et al., Extended hybrid controller design of bifurcation in a delayed chemostat model, MATCH Commun. Math. Comput. Chem., 90 (2023), 609–648. https://doi.org/10.46793/match.90-3.609X doi: 10.46793/match.90-3.609X
|
| [24] |
P. Li, X. Peng, C. Xu, L. Han, S. Shi, Novel extended mixed controller design for bifurcation control of fractional-order Myc/E2F/miR-17-92 network model concerning delay, Math. Methods Appl. Sci., 46 (2023), 18878–18898. https://doi.org/10.1002/mma.9597 doi: 10.1002/mma.9597
|
| [25] |
Y. Zhang, P. Li, C. Xu, X. Peng, R. Qiao, Investigating the effects of a fractional operator on the evolution of the enso model: bifurcations, stability and numerical analysis, Fractal Fract., 7 (2023), 602. https://doi.org/10.3390/fractalfract7080602 doi: 10.3390/fractalfract7080602
|
| [26] | K. H. Hntsa, Z. T. Mengesha, Mathematical modelling of fish resources harvesting with predator at maximum sustainable yield, Int. J. Integr. Sci. Innovation Technol., 5 (2016), 7–24. |
| [27] | J. Guckenheimer, P. Holmes, Nonlinear oscillations, dynamical systems and bifurcation of vector fields, Springer, 1983. https://doi.org/10.1007/978-1-4612-1140-2 |
| [28] | Y. A. Kuznetsov, Elements of applied bifurcation theorey, Springer, 2004. https://doi.org/10.1007/978-1-4757-3978-7 |
| [29] |
W. Liu, D. Cai, Bifurcation, chaos analysis and control in a discrete-time predator-prey system, Adv. Differ. Equations, 2019 (2019), 11. https://doi.org/10.1186/s13662-019-1950-6 doi: 10.1186/s13662-019-1950-6
|
| [30] |
Y. Liu, X. Li, Dynamics of a discrete predator-prey model with Holling-Ⅱ functional response, Int. J. Biomath., 14 (2021), 2150068. https://doi.org/10.1142/S1793524521500686 doi: 10.1142/S1793524521500686
|
| [31] |
Q. Shu, J. Xie, Stability and bifurcation analysis of discrete predator-prey model with nonlinear prey harvesting and prey refuge, Math. Methods Appl. Sci., 45 (2022), 3589–3604. https://doi.org/10.1002/mma.8005 doi: 10.1002/mma.8005
|
| [32] |
P. K. Santra, H. S. Panigoro, G. S. Mahapatra, Complexity of a discrete-time predator-prey model involving prey refuge proportional to predator, Jambura J. Math., 4 (2022), 50–63. https://doi.org/10.34312/jjom.v4i1.11918 doi: 10.34312/jjom.v4i1.11918
|
| [33] |
M. Chen, R. Wu, H. Liu, X. Fu, Spatiotemporal complexity in a Leslie-Gower type predator-prey model near Turing-Hopf point, Chaos Solitons Fract., 153 (2021), 111509. https://doi.org/10.1016/j.chaos.2021.111509 doi: 10.1016/j.chaos.2021.111509
|
| [34] |
A. Q. Khan, M. B. Javaid, Discrete-time phytoplankton-zooplankton model with bifurcations and chaos, Adv. Differ. Equations, 2021 (2021), 415. https://doi.org/10.1186/s13662-021-03523-5 doi: 10.1186/s13662-021-03523-5
|
| [35] |
A. Q. Khan, J. Ma, D. Xiao, Bifurcations of a two-dimensional discrete time plant-herbivore system, Commun. Nonlinear Sci. Numer. Simul., 39 (2016), 185–198. https://doi.org/10.1016/j.cnsns.2016.02.037 doi: 10.1016/j.cnsns.2016.02.037
|
| [36] |
A. Q. Khan, J. Ma, D. Xiao, Global dynamics and bifurcation analysis of a host-parasitoid model with strong Allee effect, J. Biol. Dyn., 11 (2017), 121–146. https://doi.org/10.1080/17513758.2016.1254287 doi: 10.1080/17513758.2016.1254287
|
| [37] |
M. Parsamanesh, M. Erfanian, Stability and bifurcations in a discrete-time SIVS model with saturated incidence rate, Chaos Solitons Fract., 150 (2021), 111178. https://doi.org/10.1016/j.chaos.2021.111178 doi: 10.1016/j.chaos.2021.111178
|
| [38] | S. N. Elaydi, An Introduction to difference equations, Springer-Verlag, 1996. https://doi.org/10.1007/0-387-27602-5 |
| [39] | S. Lynch, Dynamical systems with applications using mathematica, Birkhäuser, 2007. https://doi.org/10.1007/978-0-8176-4586-1 |
Figures(6) / Tables(1)

A. Q. Khan, Ibraheem M. Alsulami, S. K. A. Hamdani. Controlling the chaos and bifurcations of a discrete prey-predator model[J]. AIMS Mathematics, 2024, 9(1): 1783-1818. doi: 10.3934/math.2024087







 DownLoad:
DownLoad: