This paper studies the persistence, extinction and practical exponential stability of impulsive stochastic competition models with time-varying delays. The existence of the global positive solutions is investigated by the relationship between the solutions of the original system and the equivalent system, and the sufficient conditions of system persistence and extinction are given. Moreover, our study shows the following facts: (1) The impulsive perturbation does not affect the practical exponential stability under the condition of bounded pulse intensity. (2) In solving the stability of non-Markovian processes, it can be transformed into solving the stability of Markovian processes by applying Razumikhin inequality. (3) In some cases, a non-Markovian process can produce Markovian effects. Finally, numerical simulations obtained the importance and validity of the theoretical results for the existence of practical exponential stability through the relationship between parameters, pulse intensity and noise intensity.
Citation: Yuxiao Zhao, Hong Lin, Xiaoyan Qiao. Persistence, extinction and practical exponential stability of impulsive stochastic competition models with varying delays[J]. AIMS Mathematics, 2023, 8(10): 22643-22661. doi: 10.3934/math.20231152
This paper studies the persistence, extinction and practical exponential stability of impulsive stochastic competition models with time-varying delays. The existence of the global positive solutions is investigated by the relationship between the solutions of the original system and the equivalent system, and the sufficient conditions of system persistence and extinction are given. Moreover, our study shows the following facts: (1) The impulsive perturbation does not affect the practical exponential stability under the condition of bounded pulse intensity. (2) In solving the stability of non-Markovian processes, it can be transformed into solving the stability of Markovian processes by applying Razumikhin inequality. (3) In some cases, a non-Markovian process can produce Markovian effects. Finally, numerical simulations obtained the importance and validity of the theoretical results for the existence of practical exponential stability through the relationship between parameters, pulse intensity and noise intensity.

| [1] |
M. Liu, K. Wang, Persistence, extinction and global asymptotical stability of a non-autonomous predator-prey model with random perturbation, Appl. Math. Model., 36 (2012), 5344–5353. https://doi.org/10.1016/j.apm.2011.12.057 doi: 10.1016/j.apm.2011.12.057

|
| [2] |
L. Zu, D. Jiang, D. {O}'Regan, B. Ge, Periodic solution for a non-autonomous Lotka-Volterra predator-prey model with random perturbation, J. Math. Anal. Appl., 430 (2015), 428–437. https://doi.org/10.1016/j.jmaa.2015.04.058 doi: 10.1016/j.jmaa.2015.04.058
|
| [3] |
S. Zhang, X. Meng, T. Feng, T. Zhang, Dynamics analysis and numerical simulations of a stochasticnon-autonomous predator-prey system with impulsive effects, Nonlinear Anal.-Hybri., 26 (2017), 19–37. https://doi.org/10.1016/j.nahs.2017.04.003 doi: 10.1016/j.nahs.2017.04.003
|
| [4] |
S. Cheng, Stochastic population systems, Stoch. Anal. Appl., 27 (2009), 854–874. https://doi.org/10.1080/07362990902844348 doi: 10.1080/07362990902844348
|
| [5] | Z. Ma, Z. Luo, Z. Jin, H. Zhao, The threshold of survival for predator-prey volterra system of three species in a polluted environment, Syst. Sci. Math. Complex., 8 (1995), 373–382. |
| [6] |
V. Hutson, G. T. Vickers, A criterion for permanent coexistence of species with an application to a two-prey one-predator system, Math. Biosci., 63 (1983), 253–269. https://doi.org/10.1016/0025-5564(82)90042-6 doi: 10.1016/0025-5564(82)90042-6
|
| [7] |
S. Ahmad, I. M. Stamova, Almost necessary and sufficient conditions for survival of species, Nonlinear Anal.-Real., 5 (2004), 219–229. https://doi.org/10.1016/S1468-1218(03)00037-3 doi: 10.1016/S1468-1218(03)00037-3
|
| [8] |
H. I. Freedman, P. Waltman, Mathematical analysis of some three-species food chain models, Math. Biosci., 33 (1977), 257–276. https://doi.org/10.1016/0025-5564(77)90142-0 doi: 10.1016/0025-5564(77)90142-0
|
| [9] |
H. Qiu, W. Deng, Stationary distribution and global asymptotic stability of a three-species stochastic food-chain system, Turk. J. Math., 41 (2017), 5. https://doi.org/10.3906/mat-1510-52 doi: 10.3906/mat-1510-52
|
| [10] |
X. Li, J. Cao, An impulsive delay inequality involving unbounded time-varying delay and applications, IEEE T. Automat. Contr., 62 (2017), 3618–3625. https://doi.org/10.1109/TAC.2017.2669580 doi: 10.1109/TAC.2017.2669580
|
| [11] |
C. Lu, J. Chen, X. Fan, L. Zhang, Dynamics and simulations of a stochastic predator-prey model with infinite delay and impulsive perturbations, J. Appl. Math. Comput., 57 (2018), 437–465. https://doi.org/10.1007/s12190-017-1114-3 doi: 10.1007/s12190-017-1114-3
|
| [12] |
W. Hu, Q. Zhu, Stability analysis of impulsive stochastic delayed differential systems with unbounded delays, Syst. Control Lett., 136 (2020), 104606. https://doi.org/10.1016/j.sysconle.2019.104606 doi: 10.1016/j.sysconle.2019.104606
|
| [13] |
R. Rao, Z. Lin, X. Ai, J. Wu, Synchronization of epidemic systems with Neumann boundary value under delayed impulse, Mathematics, 10 (2022), 2064. https://doi.org/10.3390/math10122064 doi: 10.3390/math10122064
|
| [14] |
Q. Zhu, Stabilization of stochastic nonlinear delay systems with exogenous disturbances and the event-triggered feedback control, IEEE T. Automat Contr., 64 (2019), 3764–3771. https://doi.org/10.1109/TAC.2018.2882067 doi: 10.1109/TAC.2018.2882067
|
| [15] |
Y. Zhao, L. Wang, Practical exponential stability of impulsive stochastic food chain system with time-varying delays, Mathematics, 11 (2023), 147. https://doi.org/10.3390/math11010147 doi: 10.3390/math11010147
|
| [16] |
Q. Zhu, T. Huang, H$_\infty$ control of stochastic networked control systems with time-varying delays: The event-triggered sampling case, Int. J. Robust Nonlin., 31 (2021), 9767–9781. https://doi.org/10.1002/rnc.5798 doi: 10.1002/rnc.5798
|
| [17] |
Y. Tang, L. Zhou, J. Tang, Y. Rao, H. Fan, J. Zhu, Hybrid impulsive pinning control for mean square Synchronization of uncertain multi-link complex networks with stochastic characteristics and hybrid delays, Mathematics, 11 (2023), 1697. https://doi.org/10.3390/math11071697 doi: 10.3390/math11071697
|
| [18] |
Y. Zhao, Q. Zhu, Stabilization of stochastic highly nonlinear delay systems with neutral term, IEEE T. Automat Contr., 68 (2023), 2544–2551. https://doi.org/10.1109/TAC.2022.3186827 doi: 10.1109/TAC.2022.3186827
|
| [19] |
W. Hu, Q. Zhu, Moment exponential stability of stochastic nonlinear delay systems with impulse effects at random times, Int. J. Robust Nonlin., 29 (2019), 3809–3820. https://doi.org/10.1002/rnc.4031 doi: 10.1002/rnc.4031
|
| [20] |
M. Xia, L. Liu, J. Fang, Y. Zhang, Stability analysis for a class of stochastic differential equations with impulses, Mathematics, 11 (2023), 1541. https://doi.org/10.3390/math11061541 doi: 10.3390/math11061541
|
| [21] |
H. Xu, Q. Zhu, New criteria on $p$th moment exponential stability of stochastic delayed differential systems subject to average-delay impulses, Syst. Control Lett., 164 (2022), 105234. https://doi.org/10.1016/j.sysconle.2022.105234 doi: 10.1016/j.sysconle.2022.105234
|
| [22] |
G. Li, Y. Zhang, Y. Guan, W. Li, Stability analysis of multi-point boundary conditions for fractional differential equation with non-instantaneous integral impulse, Math. Biosci. Eng., 20 (2023), 7020–7041. https://doi.org/10.3934/mbe.2023303 doi: 10.3934/mbe.2023303
|
| [23] |
R. Wu, X. Zou, K. Wang, Asymptotic behavior of a stochastic non-autonomous predator-prey model with impulsive perturbations, Commun. Nonlinear Sci., 20 (2015), 965–974. https://doi.org/10.1016/j.cnsns.2014.06.023 doi: 10.1016/j.cnsns.2014.06.023
|
| [24] |
W. Zuo, D. Jiang, Periodic solutions for a stochastic non-autonomous Holling-Tanner predator-prey system with impulses, Nonlinear Anal.-Hybri., 22 (2016), 191–201. https://doi.org/10.1016/j.nahs.2016.03.004 doi: 10.1016/j.nahs.2016.03.004
|
| [25] |
C. Lu, X. Ding, Persistence and extinction of a stochastic logistic model with delays and impulsive perturbations, Acta Math. Sci., 34 (2014), 1551–1570. https://doi.org/10.1016/S0252-9602(14)60103-X doi: 10.1016/S0252-9602(14)60103-X
|
| [26] |
Y. Zhao, L. Wang, Y. Wang, The periodic solutions to a stochastic two-prey one-predator population model with impulsive perturbations in a polluted environment, Methodol Comput. Appl. Probab., 23 (2021), 859–872. https://doi.org/10.1007/s11009-020-09790-1 doi: 10.1007/s11009-020-09790-1
|
| [27] |
X. Fu, Q. Zhu, Stability of nonlinear impulsive stochastic systems with Markovian switching under generalized average dwell time condition, Sci. China Inf. Sci., 61 (2018), 112211. https://doi.org/10.1007/s11432-018-9496-6 doi: 10.1007/s11432-018-9496-6
|
| [28] |
T. Caraballo, M. A. Hammami, L. Mchiri, Practical asymptotic stability of nonlinear stochastic evolution equations, Stoch. Anal. Appl., 32 (2014), 77–87. https://doi.org/10.1080/07362994.2013.843142 doi: 10.1080/07362994.2013.843142
|
| [29] |
Q. Yao, P. Lin, L. Wang, Y. Wang, Practical exponential stability of impulsive stochastic Reaction-Diffusion systems with delays stochastics, IEEE T. Cybernetics, 52 (2022), 2687–2697. https://doi.org/10.1109/TCYB.2020.3022024 doi: 10.1109/TCYB.2020.3022024
|
| [30] |
T. Caraballo, M. A. Hammamib, L. Mchirib, Practical exponential stability of impulsive stochastic functional differential equations, Syst. Control Lett., 109 (2017), 43–48. https://doi.org/10.1016/j.sysconle.2017.09.009 doi: 10.1016/j.sysconle.2017.09.009
|
| [31] |
G. Tan, Z. Wang, Reachable set estimation of delayed Markovian jump neural networks based on an improved reciprocally convex inequality, IEEE T. Neur. Net. Lear., 33 (2022), 2737–2742. https://doi.org/10.1109/TNNLS.2020.3045599 doi: 10.1109/TNNLS.2020.3045599
|
| [32] |
G. Tan, Z. Wang, Stability analysis of systems with time-varying delay via a delay-product-type integral inequality, Math. Methods Appl. Sci., 45 (2022), 6535–6545. https://doi.org/10.1002/mma.8186 doi: 10.1002/mma.8186
|
| [33] |
J. Geng, M. Liu, Y. Zhang, Stability of a stochastic one-predator-two-prey population model with time delays, Commun. Nonlinear Sci., 53 (2007), 65–82. https://doi.org/10.1016/j.cnsns.2017.04.022 doi: 10.1016/j.cnsns.2017.04.022
|
| [34] |
C. Lu, X. Ding, Persistence and extinction of a stochastic logistic model with delays and impulsive perturbation, Acta Math. Sci., 34 (2014), 1551–1570. https://doi.org/10.1016/S0252-9602(14)60103-X doi: 10.1016/S0252-9602(14)60103-X
|
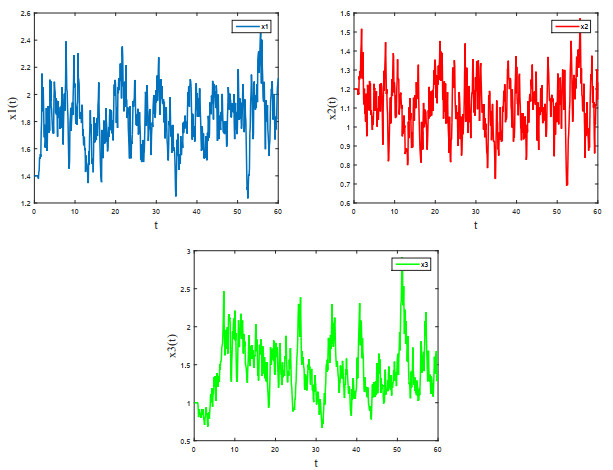
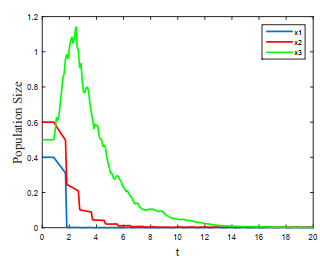
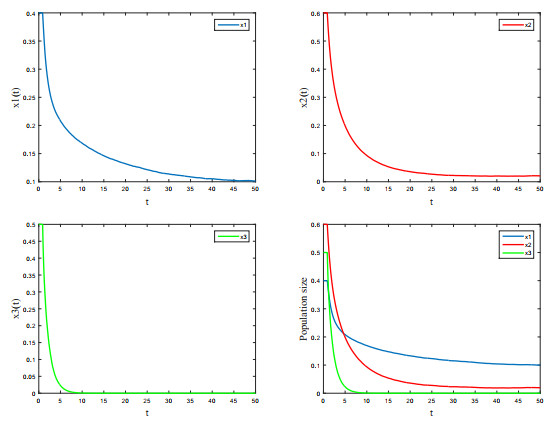
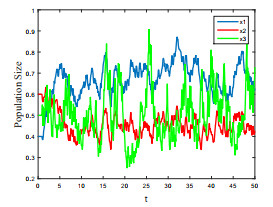
Figures(4) / Tables(1)

Yuxiao Zhao, Hong Lin, Xiaoyan Qiao. Persistence, extinction and practical exponential stability of impulsive stochastic competition models with varying delays[J]. AIMS Mathematics, 2023, 8(10): 22643-22661. doi: 10.3934/math.20231152







 DownLoad:
DownLoad: