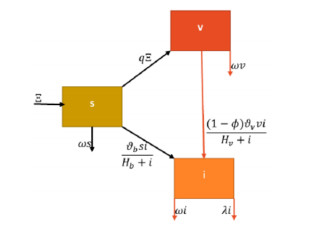
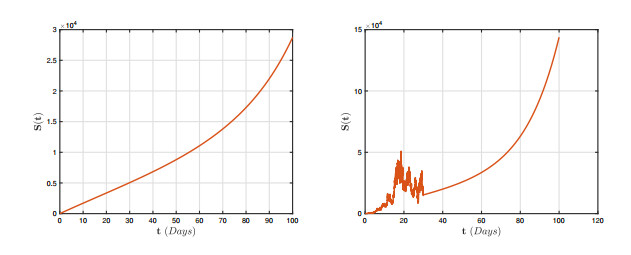
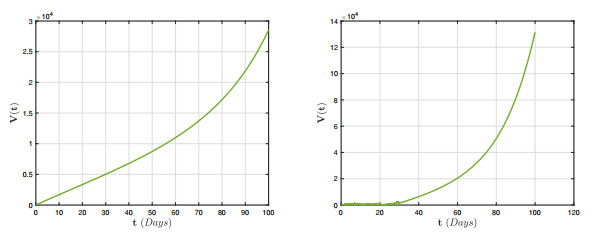
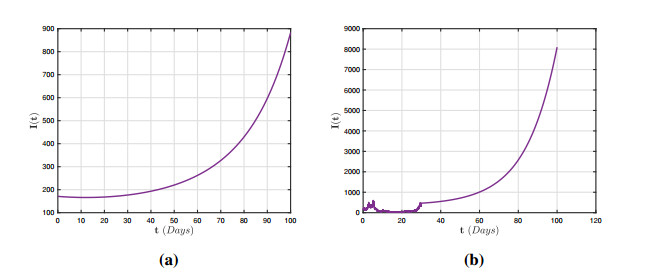
In this research, we investigate the dynamical behaviour of a HPAI epidemic system featuring a half-saturated transmission rate and significant evidence of crossover behaviours. Although simulations have proposed numerous mathematical frameworks to portray these behaviours, it is evident that their mathematical representations cannot adequately describe the crossover behaviours, particularly the change from deterministic reboots to stochastics. Furthermore, we show that the stochastic process has a threshold number $ {\bf R}_{0}^{s} $ that can predict pathogen extermination and mean persistence. Furthermore, we show that if $ {\bf R}_{0}^{s} > 1 $, an ergodic stationary distribution corresponds to the stochastic version of the aforementioned system by constructing a sequence of appropriate Lyapunov candidates. The fractional framework is expanded to the piecewise approach, and a simulation tool for interactive representation is provided. We present several illustrated findings for the system that demonstrate the utility of the piecewise estimation technique. The acquired findings offer no uncertainty that this notion is a revolutionary viewpoint that will assist mankind in identifying nature.
Citation: Maysaa Al-Qureshi, Saima Rashid, Fahd Jarad, Mohammed Shaaf Alharthi. Dynamical behavior of a stochastic highly pathogenic avian influenza A (HPAI) epidemic model via piecewise fractional differential technique[J]. AIMS Mathematics, 2023, 8(1): 1737-1756. doi: 10.3934/math.2023089
In this research, we investigate the dynamical behaviour of a HPAI epidemic system featuring a half-saturated transmission rate and significant evidence of crossover behaviours. Although simulations have proposed numerous mathematical frameworks to portray these behaviours, it is evident that their mathematical representations cannot adequately describe the crossover behaviours, particularly the change from deterministic reboots to stochastics. Furthermore, we show that the stochastic process has a threshold number $ {\bf R}_{0}^{s} $ that can predict pathogen extermination and mean persistence. Furthermore, we show that if $ {\bf R}_{0}^{s} > 1 $, an ergodic stationary distribution corresponds to the stochastic version of the aforementioned system by constructing a sequence of appropriate Lyapunov candidates. The fractional framework is expanded to the piecewise approach, and a simulation tool for interactive representation is provided. We present several illustrated findings for the system that demonstrate the utility of the piecewise estimation technique. The acquired findings offer no uncertainty that this notion is a revolutionary viewpoint that will assist mankind in identifying nature.

| [1] |
B. Joob, W. Viroj, H5N6 influenza virus infection, the newest influenza, Asian Pac. J. Trop. Bio., 5 (2015), 434–437. https://doi.org/10.1016/j.apjtb.2015.03.001 doi: 10.1016/j.apjtb.2015.03.001

|
| [2] | F. Claes, S. Von Dobschuetz, Avian influenza A(H5N6): The latest addition to emerging zoonotic avian influenza threats in east and southeast Asia, Empres Watch, 30 (2014). |
| [3] | World Health Organization, Weekly epidemiological record Relevpidmiologique hebdomadaire, 92 (2017), 453–476. Available from: http://www.who.int/wer. |
| [4] | Centers for Disease Control and Prevention, HPAI A H5 virus background and clinical illness. https://www.cdc.gov/flu/avianflu/hpai/hpai-background-clinical-illness.htm |
| [5] |
N. S. Chong, R. J. Smith, Modeling avian influenza using Filippov systems to determine culling of infected birds and quarantine, Nonlinear Anal. Real, 24 (2015), 196–218. https://doi.org/10.1016/j.nonrwa.2015.02.007 doi: 10.1016/j.nonrwa.2015.02.007
|
| [6] |
Z. Liu, C. T. Fang, A modeling study of human infections with avian influenza A H7N9 virus in mainland China, Int. J. Infect. Dis., 41 (2015), 73–78. https://doi.org/10.1016/j.ijid.2015.11.003 doi: 10.1016/j.ijid.2015.11.003
|
| [7] |
X. Zhang, L. Zou, J. Chen, Y. Fang, J. Huang, J. Zhang, et al., Avian influenza A H7N9 virus has been established in China, J. Biol. Syst., 25 (2017), 605–623. https://doi.org/10.1142/S0218339017400095 doi: 10.1142/S0218339017400095
|
| [8] | F. Brauer, C. Castillo-Chavez, Mathematical models for communicable diseases, 2012. |
| [9] |
N. S. Chong, J. M. Tchuenche, R. J. Smith, A mathematical model of avian influenza with half-saturated incidence, Theory Biosci., 133 (2014), 23–38. http://doi.org/10.1007/s12064-013-0183-6 doi: 10.1007/s12064-013-0183-6
|
| [10] |
S. Liu, S. Ruan, X. Zhang, On avian influenza epidemic models with time delay, Theory Biosci., 134 (2015), 75–82. https://doi.org/10.1007/s12064-015-0212-8 doi: 10.1007/s12064-015-0212-8
|
| [11] |
S. N. Hajiseyedazizi, M. E. Samei, J. Alzabut, Y. M. Chu, On multi-step methods for singular fractional q-integro-differential equations, Open Math., 19 (2021), 1378–1405. https://doi.org/10.1515/math-2021-0093 doi: 10.1515/math-2021-0093
|
| [12] | A. A. Kilbas, O. I. Marichev, S. G. Samko, Fractional integral and derivatives: Theory and applications, 1993. |
| [13] | K. S. Miller, B. Ross, An introduction to the fractional calculus and fractional differential equations, Wiley, 1993. |
| [14] | A. A. Kilbas, H. M. Shrivastava, J. J. Trujillo, Theory and applications of fractional differential equations, Elsevier, 2006. |
| [15] | T. H. Zhao, O. Castillo, H. Jahanshahi, A. Yusuf, M. O. Alassafi, F. E. Alsaadi, et al., A fuzzy based strategy to suppress the novel coronavirus (2019-NCOV) massive outbreak, Appl. Comput. Math., 20 (2021), 160–176. |
| [16] |
T. H. Zhao, M. I. Khan, Y. M. Chu, Artificial neural networking (ANN) analysis for heat and entropy generation in flow of non-Newtonian fluid between two rotating disks, Math. Method. Appl. Sci., 2021. https://doi.org/10.1002/mma.7310 doi: 10.1002/mma.7310
|
| [17] |
K. Karthikeyan, P. Karthikeyan, H. M. Baskonus, K. Venkatachalam, Y. M. Chu, Almost sectorial operators on $\Psi$-Hilfer derivative fractional impulsive integro-differential equations, Math. Method. Appl. Sci., 45 (2022), 8045–8059. https://doi.org/10.1002/mma.7954 doi: 10.1002/mma.7954
|
| [18] |
M. Caputo, M. Fabrizio, A new definition of fractional derivative without singular kernel, Progr. Fract. Differ. Appl., 1 (2015), 73–85. https://doi.org/10.12785/pfda/010201 doi: 10.12785/pfda/010201
|
| [19] |
A. Atangana, D. Baleanu, New fractional derivatives with non-local and nonsingular kernel: Theory and application to heat transfer model, Therm. Sci., 20 (2016), 763–769. https://doi.org/10.2298/TSCI160111018A doi: 10.2298/TSCI160111018A
|
| [20] |
Y. M. Chu, U. Nazir, M. Sohail, M. M. Selim, J. R. Lee, Enhancement in thermal energy and solute particles using hybrid nanoparticles by engaging activation energy and chemical reaction over a parabolic surface via finite element approach, Fractal Fract., 5 (2021), 119. https://doi.org/10.3390/fractalfract5030119 doi: 10.3390/fractalfract5030119
|
| [21] |
S. Rashid, S. Sultana, Y. Karaca, A. Khalid, Y. M. Chu, Some further extensions considering discrete proportional fractional operators, Fractals, 30 (2022), 2240026. https://doi.org/10.1142/S0218348X22400266 doi: 10.1142/S0218348X22400266
|
| [22] |
S. Rashid, E. I. Abouelmagd, A. Khalid, F. B. Farooq, Y. M. Chu, Some recent developments on dynamical h-discrete fractional type inequalities in the frame of nonsingular and nonlocal kernels, Fractals, 30 (2022), 2240110. https://doi.org/10.1142/S0218348X22401107 doi: 10.1142/S0218348X22401107
|
| [23] |
G. Behzad, A. Atangana, A new application of fractional Atangana–Baleanu derivatives: designing ABC-fractional masks in image processing, Phys. A, 542 (2020), 123516. https://doi.org/10.1016/j.physa.2019.123516 doi: 10.1016/j.physa.2019.123516
|
| [24] |
S. Rashid, B. Kanwal, A. G. Ahmad, E. Bonyah, S. K. Elagan, Novel numerical estimates of the pneumonia and meningitis epidemic model via the nonsingular kernel with optimal analysis, Complexity, 2022 (2022), 4717663. https://doi.org/10.1155/2022/4717663 doi: 10.1155/2022/4717663
|
| [25] |
S. Rashid, A. Khalid, S. Sultana, F. Jarad, K. M. Abualnaja, Y. S. Hamed, Novel numerical investigation of the fractional oncolytic effectiveness model with M1 virus via generalized fractional derivative with optimal criterion, Results Phys., 37 (2022), 105553. https://doi.org/10.1016/j.rinp.2022.105553 doi: 10.1016/j.rinp.2022.105553
|
| [26] |
W. Gao, H. M. Baskonus, L. Shi, New investigation of bats-hosts-reservoir-people coronavirus model and application to 2019-nCoV system, Adv. Differ. Equ., 2020 (2020), 391. https://doi.org/10.1186/s13662-020-02831-6 doi: 10.1186/s13662-020-02831-6
|
| [27] |
E. F. D. Goufo, Y. Khan, Q. A. Chaudhry, HIV and shifting epicenters for COVID-19, an alert for some countries, Chaos Soliton. Fract., 139 (2020), 110030. https://doi.org/10.1016/j.chaos.2020.110030 doi: 10.1016/j.chaos.2020.110030
|
| [28] |
A. Atangana, S. I. Araz, Modeling third waves of Covid-19 spread with piecewise differential and integral operators: Turkey, Spain and Czechia, Results Phys., 29 (2021), 104694. https://doi.org/10.1016/j.rinp.2021.104694 doi: 10.1016/j.rinp.2021.104694
|
| [29] |
M. A. Khan, H. P. Odinsyah, Fractional model of HIV transmission with awareness effect, Chaos Soliton. Fract., 138 (2020), 109967. https://doi.org/10.1016/j.chaos.2020.109967 doi: 10.1016/j.chaos.2020.109967
|
| [30] |
F. Z. Wang, M. N. Khan, I. Ahmad, H. Ahmad, H. Abu-Zinadah, Y. M. Chu, Numerical solution of traveling waves in chemical kinetics: Time-fractional fishers equations, Fractals, 30 (2022), 2240051. https://doi.org/10.1142/S0218348X22400515 doi: 10.1142/S0218348X22400515
|
| [31] |
S. Rashid, E. I. Abouelmagd, S. Sultana, Y. M. Chu, New developments in weighted n-fold type inequalities via discrete generalized h-proportional fractional operators, Fractals, 30 (2022), 2240056. https://doi.org/10.1142/S0218348X22400564 doi: 10.1142/S0218348X22400564
|
| [32] |
S. N. Hajiseyedazizi, M. E. Samei, J. Alzabut, Y. M. Chu, On multi-step methods for singular fractional q-integro-differential equations, Open Math., 19 (2021), 1378–1405. https://doi.org/10.1515/math-2021-0093 doi: 10.1515/math-2021-0093
|
| [33] |
S. A. Iqbal, M. G. Hafez, Y. M. Chu, C. Park, Dynamical analysis of nonautonomous RLC circuit with the absence and presence of Atangana-Baleanu fractional derivative, J. Appl. Anal. Comput., 12 (2022), 770–789. https://doi.org/10.11948/20210324 doi: 10.11948/20210324
|
| [34] | T. H. Zhao, O. Castillo, H. Jahanshahi, A. Yusuf, M. O. Alassafi, F. E. Alsaadi, et al., A fuzzy-based strategy to suppress the novel coronavirus (2019-NCOV) massive outbreak, Appl. Comput. Math., 20 (2021), 160–176. |
| [35] |
Y. Zhao, E. E. Elattar, M. A. Khan, Fatmawati, M. Asiri, P. Sunthrayuth, The dynamics of the HIV/AIDS infection in the framework of piecewise fractional differential equation, Results Phys., 40 (2022), 105842. https://doi.org/10.1016/j.rinp.2022.105842 doi: 10.1016/j.rinp.2022.105842
|
| [36] |
A. Atangana, S. I. Araz, Modeling and forecasting the spread of COVID-19 with stochastic and deterministic approaches: Africa and Europe, Adv. Differ. Equ., 2021 (2021), 57. https://doi.org/10.1186/s13662-021-03213-2 doi: 10.1186/s13662-021-03213-2
|
| [37] |
S. Rashid, M. K. Iqbal, A. M. Alshehri, R. Ashraf, F. Jarad, A comprehensive analysis of the stochastic fractal–fractional tuberculosis model via Mittag-Leffler kernel and white noise, Results Phys., 39 (2022), 105764. https://doi.org/10.1016/j.rinp.2022.105764 doi: 10.1016/j.rinp.2022.105764
|
| [38] |
M. Al Qurashi, S. Rashid, F. Jarad, A computational study of a stochastic fractal-fractional hepatitis B virus infection incorporating delayed immune reactions via the exponential decay, Math. Biosci. Eng., 19 (2022), 12950–12980. https://doi.org/10.3934/mbe.2022605 doi: 10.3934/mbe.2022605
|
| [39] |
S. Rashid, R. Ashraf, Q. Asif, F. Jarad, Novel dynamics of a stochastic fractal-fractional immune effector response to viral infection via latently infectious tissues, Math. Biosci. Eng., 19 (2022), 11563–11594. https://doi.org/10.3934/mbe.2022539 doi: 10.3934/mbe.2022539
|
| [40] |
B. Zhou, X. Zhang, D. Jiang, A Dynamics and density function analysis of a stochastic SVI epidemic model with half saturated incidence rate, Chaos Soliton. Fract., 137 (2020), 109865. https://doi.org/10.1016/j.chaos.2020.109865 doi: 10.1016/j.chaos.2020.109865
|
| [41] | X. Mao, Stochastic differential equations and applications, Horwood Publishing, 1997. |
| [42] |
X. Mao, G. Marion, E. Renshaw, Environmental Brownian noise suppresses explosions in population dynamics, Stoch. Proc. Appl., 97 (2002), 95–110. https://doi.org/10.1016/S0304-4149(01)00126-0 doi: 10.1016/S0304-4149(01)00126-0
|
| [43] |
R. Lipster, A strong law of large numbers for local martingales, Stochastics, 3 (1980), 217–228. https://doi.org/10.1080/17442508008833146 doi: 10.1080/17442508008833146
|
| [44] | R. Hasminiskii, Stochastic stability of differential equations, 1980. |
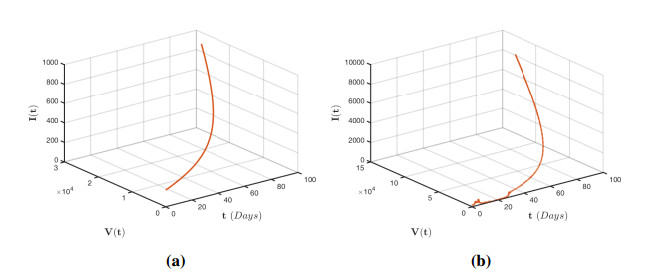
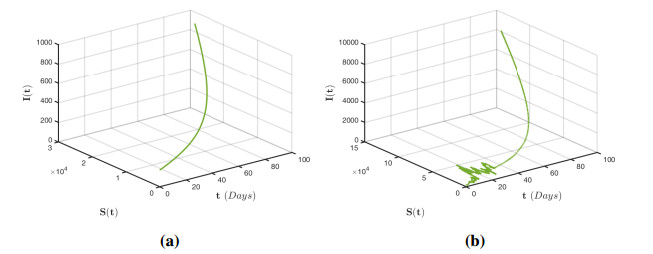
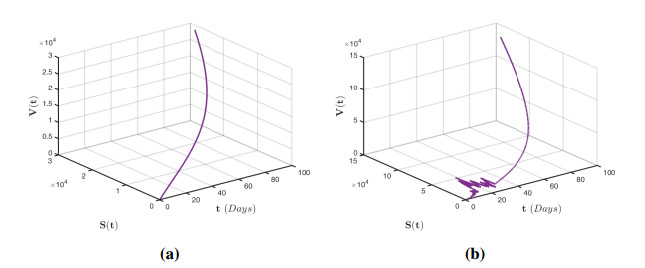
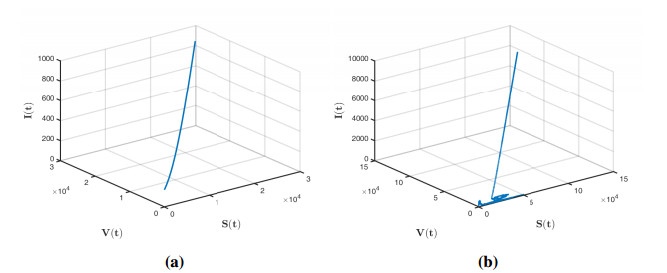
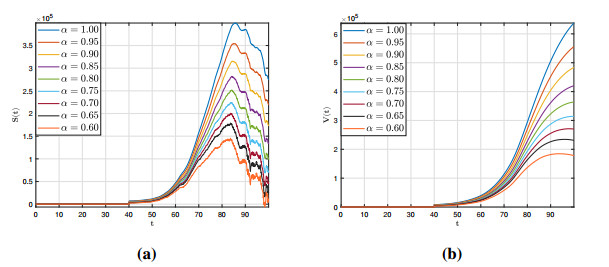
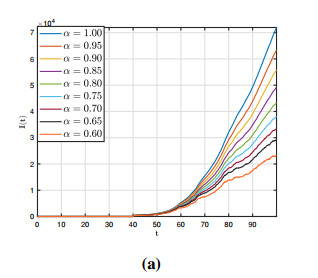
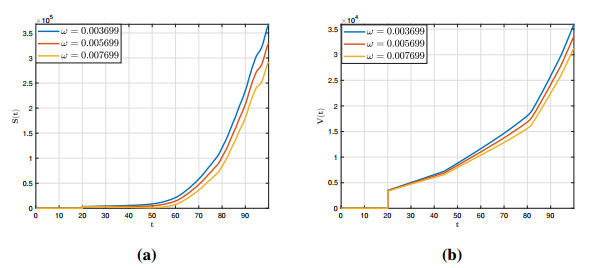
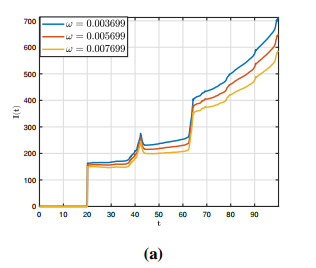
Figures(12)

Maysaa Al-Qureshi, Saima Rashid, Fahd Jarad, Mohammed Shaaf Alharthi. Dynamical behavior of a stochastic highly pathogenic avian influenza A (HPAI) epidemic model via piecewise fractional differential technique[J]. AIMS Mathematics, 2023, 8(1): 1737-1756. doi: 10.3934/math.2023089







 DownLoad:
DownLoad: