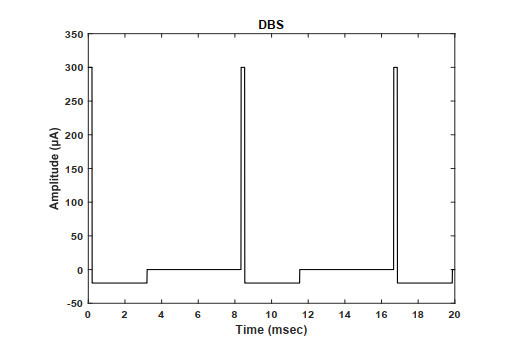
Deep brain stimulation (DBS) alleviates the symptoms of tremor, rigidity, and akinesia of the Parkinson's disease (PD). Over decades of the clinical experience, subthalamic nucleus (STN), globus pallidus externa (GPe) and globus pallidus internal (GPi) have been chosen as the common DBS target sites. However, how to design the DBS waveform is still a challenging problem. There is evidence that chronic high-frequency stimulation may cause long-term tissue damage and other side effects. In this paper, we apply a form of DBS with delayed rectangular waveform, denoted as pulse-delay-pulse (PDP) type DBS, on multiple-site based on a computational model of the basal ganglia-thalamus (BG-TH) network. We mainly investigate the effects of the stimulation frequency on relay reliability of the thalamus neurons, beta band oscillation of GPi nucleus and firing rate of the BG network. The results show that the PDP-type DBS at STN-GPe site results in better performance at lower frequencies, while the DBS at GPi-GPe site causes the number of spikes of STN to decline and deviate from the healthy status. Fairly good therapeutic effects can be achieved by PDP-type DBS at STN-GPi site only at higher frequencies. Thus, it is concluded that the application of multiple-site stimulation with PDP-type DBS at STN-GPe is of great significance in treating symptoms of neurological disorders in PD.
Citation: Xia Shi, Ziheng Zhang. Multiple-site deep brain stimulation with delayed rectangular waveforms for Parkinson's disease[J]. Electronic Research Archive, 2021, 29(5): 3471-3487. doi: 10.3934/era.2021048
Deep brain stimulation (DBS) alleviates the symptoms of tremor, rigidity, and akinesia of the Parkinson's disease (PD). Over decades of the clinical experience, subthalamic nucleus (STN), globus pallidus externa (GPe) and globus pallidus internal (GPi) have been chosen as the common DBS target sites. However, how to design the DBS waveform is still a challenging problem. There is evidence that chronic high-frequency stimulation may cause long-term tissue damage and other side effects. In this paper, we apply a form of DBS with delayed rectangular waveform, denoted as pulse-delay-pulse (PDP) type DBS, on multiple-site based on a computational model of the basal ganglia-thalamus (BG-TH) network. We mainly investigate the effects of the stimulation frequency on relay reliability of the thalamus neurons, beta band oscillation of GPi nucleus and firing rate of the BG network. The results show that the PDP-type DBS at STN-GPe site results in better performance at lower frequencies, while the DBS at GPi-GPe site causes the number of spikes of STN to decline and deviate from the healthy status. Fairly good therapeutic effects can be achieved by PDP-type DBS at STN-GPi site only at higher frequencies. Thus, it is concluded that the application of multiple-site stimulation with PDP-type DBS at STN-GPe is of great significance in treating symptoms of neurological disorders in PD.

| [1] |
M. Stephanie et al., Clinical subtypes of Parkinson's disease, Movement Disorders, 26 (2011), 51-58. doi: 10.1002/mds.23346

|
| [2] |
Akineto-rigid vs. tremor syndromes in Parkinsonism. Current Opinion in Neurology (2009) 22: 387-393.
|
| [3] |
Pathological synchronization in Parkinson's disease: Networks, models and treatments. Trends in Neurosciences (2007) 30: 357-364.
|
| [4] |
P. Choongseok, R. M. Worth and L. L. Rubchinsky, Neural dynamics in Parkinsonian brain: The boundary between synchronized and nonsynchronized dynamics, Physical Review E, 83 (2011), 042901. doi: 10.1103/PhysRevE.83.042901
|
| [5] |
What do the basal ganglia do? A modeling perspective. Biol. Cybernet. (2010) 103: 237-253.
|
| [6] |
Computational models of basal ganglia dysfunction: The dynamics is in the details. Current Opinion in Neurobiology (2017) 46: 127-135.
|
| [7] |
A. L. Benabid et al., Functional neurosurgery for movement disorders: A historical perspective, Progress in Brain Research, 175 (2009), 379-391. doi: 10.1016/S0079-6123(09)17525-8
|
| [8] | M. Astrom, Modelling, simulation, and visualization of deep Brain stimulation, [Ph.D. thesis], Linkoping University, Linkoping, Sweden, (2011). |
| [9] |
H. Marwan, My 25 stimulating years with DBS in Parkinson's disease, Journal of Parkinson's Disease, 7 (2017), S33–S41. doi: 10.3233/JPD-179007
|
| [10] |
Deep Brain Stimulation. Intervent Neurol (2012) 1: 200-212.
|
| [11] |
J. C. Lilly et al., Brief, Noninjurious Electric Waveform for Stimulation of the Brain, Science, 121 (1955), 468-469. doi: 10.2307/1681665
|
| [12] |
X. F. Wei and W. M. Grill, Impedance characteristics of deep brain stimulation electrodes in vitro and in vivo, Journal of Neural Engineering, 6 (2009), 046008. doi: 10.1088/1741-2560/6/4/046008
|
| [13] |
L. Hofmann et al., Modified pulse shapes for effective neural stimulation, Front Neuroeng, 4 (2011), 1. doi: 10.3389/fneng.2011.00009
|
| [14] |
Multi-site stimulation of subthalamic nucleus diminishes thalamocortical relay errors in a biophysical network model. Neural Netw. (2011) 24: 602-616.
|
| [15] |
O. V. Popovych et al., Pulsatile desynchronizing delayed feedback for closed-loop deep brain stimulation, PLoS ONE, 3 (2017), e0173363. doi: 10.1371/journal.pone.0173363
|
| [16] |
O. V. Popovych and P. A. Tass, Adaptive delivery of continuous and delayed feedback deep brain stimulation - a computational study, Scientific Reports, 9 (2019), 10585. doi: 10.1038/s41598-019-47036-4
|
| [17] |
Relative contributions of local cell and passing fiber activation and silencing to changes in thalamic fidelity during deep brain stimulation and lesioning: a computational modeling study. Journal of Computational Neuroscience (2012) 32: 499-519.
|
| [18] |
Y. Smith et al., Microcircuitry of the direct and indirect pathways of the basal ganglia, Neuroscience, 86 (1998), 353-387. doi: 10.1016/s0306-4522(98)00004-9
|
| [19] |
D. Terman et al., Activity patterns in a model for the subthalamopallidal network of the basal ganglia, The Journal of Neuroence, 22 (2002), 2963-2976. doi: 10.1523/jneurosci.22-07-02963.2002
|
| [20] |
High frequency stimulation of the subthalamic nucleus eliminates pathological thalamic rhythmicity in a computational model. Journal of Computational Neuroscience (2004) 16: 211-235.
|
| [21] |
H. Daniel et al., The effects of electrode material, charge density and stimulation duration on the safety of high-frequency stimulation of the subthalamic nucleus in rats, Journal of Neuroscience Methods, 138 (2004), 207-216. doi: 10.1016/j.jneumeth.2004.04.019
|
| [22] |
M. C. Rodriguez-Oroz et al., Bilateral deep brain stimulation in Parkinson's disease: A multicentre study with 4 years follow-up, Brain, 128 (2005), 2240-2249. doi: 10.1093/brain/awh571
|
| [23] |
A. A. Kuhn et al., High-frequency stimulation of the subthalamic nucleus suppresses oscillatory activity in patients with Parkinson's disease in parallel with improvement in motor
performance, Journal of Neuroscience, 28 (2008), 6165-6173. doi: 10.1523/JNEUROSCI.0282-08.2008
|
| [24] |
S. Fei et al., Model-based evaluation of closed-loop deep brain stimulation controller to adapt to dynamic changes in reference signal, Frontiers in Neuroscience, 13 (2019), 956. doi: 10.3389/fnins.2019.00956
|
| [25] |
A biophysical model of the cortex-basal ganglia-thalamus network in the 6-OHDA lesioned rat model of Parkinson's disease. Journal of Computational Neuroscience (2016) 40: 207-229.
|
| [26] |
F. Steigerwald et al., Neuronal activity of the human subthalamic nucleus in the parkinsonian
and nonparkinsonian state, Journal of Neurophysiology, 100 (2008), 2515-2524. doi: 10.1152/jn.90574.2008
|












Figures(13) / Tables(1)
Xia Shi, Ziheng Zhang. Multiple-site deep brain stimulation with delayed rectangular waveforms for Parkinson's disease[J]. Electronic Research Archive, 2021, 29(5): 3471-3487. doi: 10.3934/era.2021048







 DownLoad:
DownLoad: