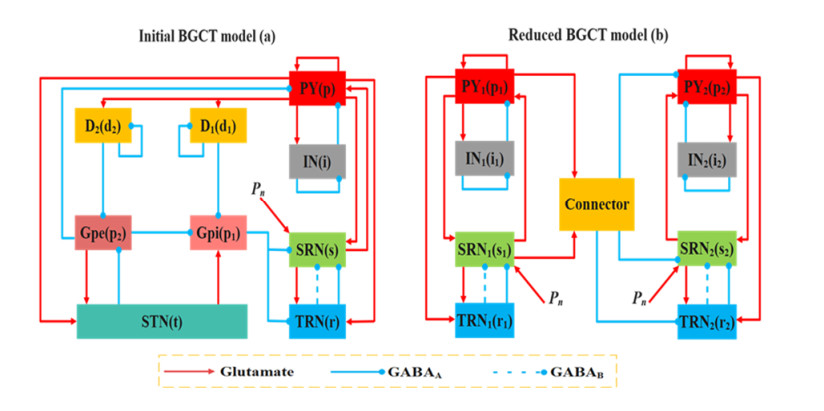
The important role of basal ganglia in corticothalamic loops has received widespread attention. However, its connection between coupled reduced corticothalamic networks is rarely researched, particularly the regulatory mechanism about electromagnetic radiation and electrical stimulation has not been comprehensively investigated. In this paper, we establish a model simplified the basal-ganglia as a connector connecting two corticothalamic loops. Four kinds of treatment methods are applied to the coupled reduced corticothalamic model, for instance deep brain stimulation (DBS), 1:0 coordinate reset stimulation (CRS) and 3:2 CRS to stimulate thalamic reticular nucleus (TRN) and electromagnetic radiation to stimulate the pyramidal neuronal population (PY). One of the important results is that the epileptic area can be significantly reduced in varying degrees by changing the strength of the basal-ganglia connector. Another one is that electromagnetic radiation, DBS and CRS have preferable inhibitory effects on absence seizure. The results show that DBS has a more significant inhibitory effect than 1:0 CRS and 3:2 CRS. The results might contribute to understanding the role of basal ganglia in coupled model and providing a reference for inhibiting epileptic seizures.
Citation: Xiaolong Tan, Hudong Zhang, Yan Xie, Yuan Chai. Electromagnetic radiation and electrical stimulation controls of absence seizures in a coupled reduced corticothalamic model[J]. Electronic Research Archive, 2023, 31(1): 58-74. doi: 10.3934/era.2023004
The important role of basal ganglia in corticothalamic loops has received widespread attention. However, its connection between coupled reduced corticothalamic networks is rarely researched, particularly the regulatory mechanism about electromagnetic radiation and electrical stimulation has not been comprehensively investigated. In this paper, we establish a model simplified the basal-ganglia as a connector connecting two corticothalamic loops. Four kinds of treatment methods are applied to the coupled reduced corticothalamic model, for instance deep brain stimulation (DBS), 1:0 coordinate reset stimulation (CRS) and 3:2 CRS to stimulate thalamic reticular nucleus (TRN) and electromagnetic radiation to stimulate the pyramidal neuronal population (PY). One of the important results is that the epileptic area can be significantly reduced in varying degrees by changing the strength of the basal-ganglia connector. Another one is that electromagnetic radiation, DBS and CRS have preferable inhibitory effects on absence seizure. The results show that DBS has a more significant inhibitory effect than 1:0 CRS and 3:2 CRS. The results might contribute to understanding the role of basal ganglia in coupled model and providing a reference for inhibiting epileptic seizures.

| [1] | R. M. Durón, M. T. Medina, I. E. Martínez-Juárez, Seizures of idiopathic generalized epilepsies, Epilepsia, 46 (2010), 34–47. https://doi.org/10.1111/j.1528-1167.2005.00312.x |
| [2] |
A. Tolaymat, A. Nayak, J. D. Geyer, Diagnosis and management of childhood epilepsy, Curr. Probl. Pediatr. Adolesc. Health Care, 45 (2015), 3–17. https://doi.org/10.1016/j.cppeds.2014.12.002 doi: 10.1016/j.cppeds.2014.12.002

|
| [3] |
V. Crunelli, N. Leresche, Childhood absence epilepsy: Genes, channels, neurons and networks, Nat. Rev. Neurosci., 3 (2002), 371–382. https://doi.org/10.1038/nrn811 doi: 10.1038/nrn811
|
| [4] |
R. Guerrini, F. Melani, C. Brancati, A. R. Ferrari, Dysgraphia as a mild expression of dystonia in children with absence epilepsy, Plos One, 10 (2015), e0130883. https://doi.org/10.1371/journal.pone.0130883 doi: 10.1371/journal.pone.0130883
|
| [5] |
E. Murphy, K. Hoshi, A. Benítez-Burraco, Subcortical syntax: Reconsidering the neural dynamics of language, J. Neurolinguistics, 62 (2022), 101062. https://doi.org/10.1016/j.jneuroling.2022.101062 doi: 10.1016/j.jneuroling.2022.101062
|
| [6] |
G. Leisman, O. Braun-Benjamin, R. Melillo, Cognitive-motor interactions of the basal ganglia in development, Front. Syst. Neurosci., 8 (2014), 16. https://doi.org/10.3389/fnsys.2014.00016 doi: 10.3389/fnsys.2014.00016
|
| [7] |
C. Marescaux, M. Vergnes, Genetic absence epilepsy in rats from strasbourg (GAERS), Ital. J. Neurol. Sci., 16 (1995), 113–118. https://doi.org/10.1007/BF02229083 doi: 10.1007/BF02229083
|
| [8] |
A. M. L. Coenen, E. L. J. M. Luijtelaar, Genetic animal models for absence epilepsy: A review of the WAG/Rij strain of rats, Behav. Genet., 33 (2003), 635–655. https://doi.org/10.1023/A:1026179013847 doi: 10.1023/A:1026179013847
|
| [9] |
I. Timofeev, M. Steriade, Neocortical seizures: Initiation, development and cessation, Neuroscience, 123 (2004), 299–336. https://doi.org/10.1016/j.neuroscience.2003.08.051 doi: 10.1016/j.neuroscience.2003.08.051
|
| [10] |
R. L. Albin, A. B. Young, J. B. Penney, The functional anatomy of basal ganglia disorders, Trends Neurosci., 12 (1989), 366–375. https://doi.org/10.1016/0166-2236(89)90074-X doi: 10.1016/0166-2236(89)90074-X
|
| [11] |
A. Parent, L. N. Hazrati, Functional anatomy of the basal ganglia. I. The cortico-basal ganglia-thalamo-cortical loop, Brain Res. Rev., 20 (1995), 91–127. https://doi.org/10.1016/0165-0173(94)00007-C doi: 10.1016/0165-0173(94)00007-C
|
| [12] |
D. Pinault, The thalamic reticular nucleus: Structure, function and concept, Brain Res. Rev., 46 (2004), 1–31. https://doi.org/10.1016/j.brainresrev.2004.04.008 doi: 10.1016/j.brainresrev.2004.04.008
|
| [13] |
R. W. Guillery, J. K. Harting, Structure and connections of the thalamic reticular nucleus: Advancing views over half a century, J. Comp. Neurol., 463 (2003), 360–371. https://doi.org/10.1002/cne.10738 doi: 10.1002/cne.10738
|
| [14] |
Z. Wang, Q. Wang, Stimulation strategies for absence seizures: Targeted therapy of the focus in coupled thalamocortical model, Nonlinear Dyn., 96 (2019), 1649–1663. https://doi.org/10.1007/s11071-019-04876-z doi: 10.1007/s11071-019-04876-z
|
| [15] |
T. Bessaïh, L. Bourgeais, C. I. Badiu, D. A. Carter, T. I. Toth, Nucleus-specific abnormalities of GABAergic synaptic transmission in a genetic model of absence seizures, J. Neurophysiol., 96 (2006), 3074–3081. https://doi.org/10.1152/jn.00682.2006 doi: 10.1152/jn.00682.2006
|
| [16] |
S. Seo, B. Leitch, Synaptic changes in GABAA receptor expression in the thalamus of the stargazer mouse model of absence epilepsy, Neuroscience, 306 (2015), 28–38. https://doi.org/10.1016/j.neuroscience.2015.08.021 doi: 10.1016/j.neuroscience.2015.08.021
|
| [17] | G. Avanzini, M. D. Curtis, C. Marescaux, F. Panzia, R. Sreafico, M. Vergnes, Role of the thalamic reticular nucleus in the generation of rhythmic thalamo-cortical activities subserving spike and waves, in Generalized Non-Convulsive Epilepsy: Focus on GABA-B Receptors, Springer, Vienna, 35 (1992), 85–95. https://doi.org/10.1007/978-3-7091-9206-1_6 |
| [18] |
Z. Nanobashvili, T. Chachua, A. Nanobashvili, I. Bilanishvili, O. Lindvall, Z. Kokaia, Suppression of limbic motor seizures by electrical stimulation in thalamic reticular nucleus, Exp. Neurol., 181 (2003), 224–230. https://doi.org/10.1016/S0014-4886(03)00045-1 doi: 10.1016/S0014-4886(03)00045-1
|
| [19] | C. R. Pantoja-Jiménez, V. M. Magdaleno-Madrigal, S. Almazán-Alvarado, R. Fernández-Masa, Anti-epileptogenic effect of high-frequency stimulation in the thalamic reticular nucleus on PTZ-induced seizures, Brain Stimul., 7 (2014), 587–594. https://doi.org/10.1016/j.brs.2014.03.012 |
| [20] |
A. Clemente-Perez, S. R. Makinson, B. Higashikubo, S. Brovarney, Distinct thalamic reticular cell types differentially modulate normal and pathological cortical rhythms, Cell Rep., 19 (2017), 2130–2142. https://doi.org/10.1016/j.celrep.2017.05.044 doi: 10.1016/j.celrep.2017.05.044
|
| [21] |
W. J. Chang, W. P. Chang, B. C. Shyu, Suppression of cortical seizures by optic stimulation of the reticular thalamus in PV-mhChR2-YFP BAC transgenic mice, Mol. Brain, 10 (2017), 1–15. https://doi.org/10.1186/s13041-017-0320-0 doi: 10.1186/s13041-017-0320-0
|
| [22] |
P. Kwan, M. J. Brodie, Early identification of refractory epilepsy, New Engl. J. Med., 342 (2000), 314–319. https://doi.org/10.1056/NEJM200002033420503 doi: 10.1056/NEJM200002033420503
|
| [23] | J. Engel, S. Wiebe, J. French, M. Sperling, P. Williamson, D. Spencer, et al., Practice parameter: temporal lobe and localized neocortical resections for epilepsy, Neurology, 60 (2003), 538–547. https://doi.org/10.1212/01.WNL.0000055086.35806.2D |
| [24] | P. Kwan, S. C. Schachter, M. J. Brodie, Drug-resistant epilepsy, New Engl. J. Med., 365 (2011), 919–926. https://doi.org/10.1056/NEJMra1004418 |
| [25] |
K. Lehtimäki, J. W. Långsjö, J. Ollikainen, H. Heinonen, M. Möttönen, Successful management of super-refractory status epilepticus with thalamic deep brain stimulation, Ann. Neurol., 81 (2017), 142–146. https://doi.org/10.1002/ana.24821 doi: 10.1002/ana.24821
|
| [26] |
I. Adamchic, C. Hauptmann, U. B. Barnikol, N. Pawelczyk, O. Popovych, T. T. Barnikol, Coordinated reset neuromodulation for Parkinson's disease: Proof-of-concept study, Mov. Disord., 29 (2014), 1679–1684. https://doi.org/10.1002/mds.25923 doi: 10.1002/mds.25923
|
| [27] |
D. Fan, Q. Wang, Closed-loop control of absence seizures inspired by feedback modulation of basal ganglia to the corticothalamic circuit, IEEE Trans. Neural Syst. Rehabil. Eng., 28 (2020), 581–590. https://doi.org/10.1109/TNSRE.2020.2969426 doi: 10.1109/TNSRE.2020.2969426
|
| [28] |
S. Majhi, D. Ghosh, Alternating chimeras in networks of ephaptically coupled bursting neurons, Chaos: An Interdiscip. J. Nonlinear Sci., 28 (2018), 083113. https://doi.org/10.1063/1.5022612 doi: 10.1063/1.5022612
|
| [29] | B. K. Bera, S. Rakshit, D. Ghosh, J. Kurths, Spike chimera states and firing regularities in neuronal hypernetworks, Chaos: An Interdiscip. J. Nonlinear Sci., 29 (2019), 053115. |
| [30] |
S. Rakshit, A. Ray, B. K. Bera, D. Ghosh, Synchronization and firing patterns of coupled Rulkov neuronal map, Nonlinear Dyn., 94 (2018), 785–805. https://doi.org/10.1007/s11071-018-4394-8 doi: 10.1007/s11071-018-4394-8
|
| [31] |
C. Wang, J. Tang, J. Ma, Minireview on signal exchange between nonlinear circuits and neurons via field coupling, Eur. Phys. J. Spec. Top., 228 (2019), 1907–1924. https://doi.org/10.1140/epjst/e2019-800193-8 doi: 10.1140/epjst/e2019-800193-8
|
| [32] |
M. Vinaya, R. P. Ignatius, Electromagnetic radiation from memristor applied to basal ganglia helps in controlling absence seizures, Nonlinear Dyn., 101 (2020), 2369–2380. https://doi.org/10.1007/s11071-020-05955-2 doi: 10.1007/s11071-020-05955-2
|
| [33] |
M. Lv, J. Ma, Multiple modes of electrical activities in a new neuron model under electromagnetic radiation, Neurocomputing, 205 (2016), 375–381. https://doi.org/10.1016/j.neucom.2016.05.004 doi: 10.1016/j.neucom.2016.05.004
|
| [34] |
M. Lv, C. Wang, G. Ren, J. Ma, X. Song, Model of electrical activity in a neuron under magnetic flow effect, Nonlinear Dyn., 85 (2016), 1479–1490. https://doi.org/10.1007/s11071-016-2773-6 doi: 10.1007/s11071-016-2773-6
|
| [35] | M. Chen, D. Guo, T. Wang, W. Jing, Y. Xia, P. Xu, et al., Bidirectional control of absence seizures by the basal ganglia: A computational evidence, PLoS Comput. Biol., 10 (2014), e1003495. https://doi.org/10.1371/journal.pcbi.1003495 |
| [36] | M. Chen, D. Guo, M. Li, S. Wu, J. Ma, Y. Cui, et al., Critical roles of the direct GABAergic pallido-cortical pathway in controlling absence seizures, PLoS Comput. Biol., 11 (2015), e1004539. https://doi.org/10.1371/journal.pcbi.1004539 |
| [37] |
P. A. Robinson, C. J. Rennie, D. L. Rowe, Dynamics of large-scale brain activity in normal arousal states and epileptic seizures, Phys. Rev. E, 65 (2002), 041924. https://doi.org/10.1103/PhysRevE.65.041924 doi: 10.1103/PhysRevE.65.041924
|
| [38] |
M. Breakspear, J. A. Roberts, J. R. Terry, S. Rodrigues, N. Mahant, P. A. Robinson, A unifying explanation of primary generalized seizures through nonlinear brain modeling and bifurcation analysis, Cereb. Cortex, 16 (2006), 1296–1313. https://doi.org/10.1093/cercor/bhj072 doi: 10.1093/cercor/bhj072
|
| [39] |
P. A. Robinson, C. J. Rennie, J. J. Wright, Propagation and stability of waves of electrical activity in the cerebral cortex, Phys. Rev. E, 56 (1997), 826–840. https://doi.org/10.1103/PhysRevE.56.826 doi: 10.1103/PhysRevE.56.826
|
| [40] |
P. A. Robinson, C. J. Rennie, J. J. Wright, H. Bahramali, E. Gordon, D. L. Rowe, Prediction of electroencephalographic spectra from neurophysiology, Phys. Rev. E, 63 (2001), 021903. https://doi.org/10.1103/PhysRevE.63.021903 doi: 10.1103/PhysRevE.63.021903
|
| [41] |
S. J. van Albada, R. T. Gray, P. M. Drysdale, P. A. Robinson, Mean-field modeling of the basal ganglia-thalamocortical system. II: Dynamics of parkinsonian oscillations, J. Theor. Biol., 257 (2009), 664–688. https://doi.org/10.1016/j.jtbi.2008.12.013 doi: 10.1016/j.jtbi.2008.12.013
|
| [42] |
B. Hu, Q. Wang, Controlling absence seizures by deep brain stimulus applied on substantia nigra pars reticulata and cortex, Chaos, Solitons Fractals, 80 (2015), 13–23. https://doi.org/10.1016/j.chaos.2015.02.014 doi: 10.1016/j.chaos.2015.02.014
|
| [43] |
M. I. Gulcebi, S. Ketenci, R. Linke, H. Hacıoğl, Topographical connections of the substantia nigra pars reticulata to higher-order thalamic nuclei in the rat, Brain Res. Bull., 87 (2012), 312–318. https://doi.org/10.1016/j.brainresbull.2011.11.005 doi: 10.1016/j.brainresbull.2011.11.005
|
| [44] |
G. D. Chiara, M. L. Porceddu, M. Morelli, M. L. Mulas, G. L. Gessa, Evidence for a GABAergic projection from the substantia nigra to the ventromedial thalamus and to the superior colliculus of the rat, Brain Res., 176 (1979), 273–284. https://doi.org/10.1016/0006-8993(79)90983-1 doi: 10.1016/0006-8993(79)90983-1
|
| [45] | P. Suffczynski, S. Kalitzin, F. L. Da Silva, Dynamics of non-convulsive epileptic phenomena modeled by a bistable neuronal network, Neuroscience, 126 (2004), 467–484. https://doi.org/10.1016/j.neuroscience.2004.03.014 |
| [46] |
Q. Li, S. Tang, H. Zeng, T. Zhou, On hyperchaos in a small memristive neural network, Nonlinear Dyn., 78 (2014), 1087–1099. https://doi.org/10.1007/s11071-014-1498-7 doi: 10.1007/s11071-014-1498-7
|
| [47] |
Q. Li, H. Zeng, J. Li, Hyperchaos in a 4D memristive circuit with infinitely many stable equilibria, Nonlinear Dyn., 79 (2015), 2295–2308. https://doi.org/10.1007/s11071-014-1812-4 doi: 10.1007/s11071-014-1812-4
|
| [48] |
D. Cafagna, G. Grassi, On the simplest fractional-order memristor-based chaotic system, Nonlinear Dynam., 70 (2012), 1185–1197. https://doi.org/10.1007/s11071-012-0522-z doi: 10.1007/s11071-012-0522-z
|
| [49] |
F. Marten, S. Rodrigues, O. Benjamin, M. P. Richardson, J. R. Terry, Onset of polyspike complexes in a mean-field model of human electroencephalography and its application to absence epilepsy, Philos. Trans. R. Soc. A, 367 (2009), 1145–1161. https://doi.org/10.1098/rsta.2008.0255 doi: 10.1098/rsta.2008.0255
|
| [50] |
R. L. Macdonald, J. Q. Kang, M. J. Gallagher, H. J. Feng, GABAA receptor mutations epilepsy associated with generalized epilepsies, Adv. Pharmacol., 54 (2006), 147–169. https://doi.org/10.1016/S1054-3589(06)54007-4 doi: 10.1016/S1054-3589(06)54007-4
|
| [51] |
A. Destexhe, Spike-and-wave oscillations based on the properties of GABAB receptors, J. Neurosci., 18 (1998), 9099–9111. https://doi.org/10.1523/JNEUROSCI.18-21-09099.1998 doi: 10.1523/JNEUROSCI.18-21-09099.1998
|







Figures(8) / Tables(2)
Xiaolong Tan, Hudong Zhang, Yan Xie, Yuan Chai. Electromagnetic radiation and electrical stimulation controls of absence seizures in a coupled reduced corticothalamic model[J]. Electronic Research Archive, 2023, 31(1): 58-74. doi: 10.3934/era.2023004







 DownLoad:
DownLoad: