Citation: Disha Mohan Bangalore, Ingrid Tessmer. Unique insight into protein-DNA interactions from single molecule atomic force microscopy[J]. AIMS Biophysics, 2018, 5(3): 194-216. doi: 10.3934/biophy.2018.3.194

| [1] |
Binnig G, Quate CF, Gerber C (1986) Atomic Force Microscope. Phys Rev Lett 56: 930–933. doi: 10.1103/PhysRevLett.56.930

|
| [2] |
Tessmer I, Kaur P, Lin JG, et al. (2013) Investigating bioconjugation by atomic force microscopy. J Nanobiotechnol 11: 1–17. doi: 10.1186/1477-3155-11-1
|
| [3] |
Werten PJL, Remigy HW, de Groot BL, et al. (2002) Progress in the analysis of membrane protein structure and function. FEBS Lett 529: 65–72. doi: 10.1016/S0014-5793(02)03290-8
|
| [4] |
Muller DJ, Sapra KT, Scheuring S, et al. (2006) Single-molecule studies of membrane proteins. Curr Opin Struct Biol 16: 489–495. doi: 10.1016/j.sbi.2006.06.001
|
| [5] |
Gorle S, Pan YG, Sun ZQ, et al. (2017) Computational Model and Dynamics of Monomeric Full-Length APOBEC3G. ACS Cent Sci 3: 1180–1188. doi: 10.1021/acscentsci.7b00346
|
| [6] | Sander B, Tria G, Shkumatov AV, et al. (2013) Structural characterization of gephyrin by AFM and SAXS reveals a mixture of compact and extended states. Acta Crystallogr 69: 2050–2060. |
| [7] |
Ishino S, Yamagami T, Kitamura M, et al. (2014) Multiple interactions of the intrinsically disordered region between the helicase and nuclease domains of the archaeal Hef protein. J Biol Chem 289: 21627–21639. doi: 10.1074/jbc.M114.554998
|
| [8] |
Shinozaki Y, Sumitomo K, Tsuda M, et al. (2009) Direct Observation of ATP-Induced Conformational Changes in Single P2X(4) Receptors. PLos Biol 7: e1000103. doi: 10.1371/journal.pbio.1000103
|
| [9] |
Lemaire PA, Tessmer I, Craig R, et al. (2006) Unactivated PKR exists in an open conformation capable of binding nucleotides. Biochemistry 45: 9074–9084. doi: 10.1021/bi060567d
|
| [10] |
Kinoshita E, van Rossum-Fikkert S, Sanchez H, et al. (2015) Human RAD50 makes a functional DNA-binding complex. Biochimie 113: 47–53. doi: 10.1016/j.biochi.2015.03.017
|
| [11] |
Bonazza K, Rottensteiner H, Seyfried BK, et al. (2014) Visualization of a protein-protein interaction at a single-molecule level by atomic force microscopy. Anal Bioanal Chem 406: 1411–1421. doi: 10.1007/s00216-013-7563-0
|
| [12] |
Shlyakhtenko LS, Gall AA, Filonov A, et al. (2003) Silatrane-based surface chemistry for immobilization of DNA, protein-DNA complexes and other biological materials. Ultramicroscopy 97: 279–287. doi: 10.1016/S0304-3991(03)00053-6
|
| [13] |
Scheuring S, Muller DJ, Ringler P, et al. (1999) Imaging streptavidin 2D crystals on biotinylated lipid monolayers at high resolution with the atomic force microscope. J Microsc 193: 28–35. doi: 10.1046/j.1365-2818.1999.00434.x
|
| [14] |
Whited AM, Park PS (2014) Atomic force microscopy: A multifaceted tool to study membrane proteins and their interactions with ligands. Biochim Biophys Acta 1838: 56–68. doi: 10.1016/j.bbamem.2013.04.011
|
| [15] |
Dubrovin EV, Schachtele M, Klinov DV, et al. (2017) Time-Lapse Single-Biomolecule Atomic Force Microscopy Investigation on Modified Graphite in Solution. Langmuir 33: 10027–10034. doi: 10.1021/acs.langmuir.7b02220
|
| [16] |
Oliveira Brett AM, Chiorcea Paquim AM (2005) DNA imaged on a HOPG electrode surface by AFM with controlled potential. Bioelectrochemistry 66: 117–124. doi: 10.1016/j.bioelechem.2004.05.009
|
| [17] |
Garcia R, Perez R (2002) Dynamic atomic force microscopy methods. Surf Sci Rep 47: 197–301. doi: 10.1016/S0167-5729(02)00077-8
|
| [18] |
Dufrene YF, Ando T, Garcia R, et al. (2017) Imaging modes of atomic force microscopy for application in molecular and cell biology. Nat Nanotechnol 12: 295–307. doi: 10.1038/nnano.2017.45
|
| [19] |
Hansma HG, Sinsheimer RL, Groppe J, et al. (1993) Recent Advances in Atomic-Force Microscopy of DNA. Scanning 15: 296–299. doi: 10.1002/sca.4950150509
|
| [20] |
Balamurugan S, Obubuafo A, Soper SA, et al. (2008) Surface immobilization methods for aptamer diagnostic applications. Anal Bioanal Chem 390: 1009–1021. doi: 10.1007/s00216-007-1587-2
|
| [21] |
Knecht S, Ricklin D, Eberle AN, et al. (2009) Oligohis-tags: Mechanisms of binding to Ni2+-NTA surfaces. J Mol Recognit 22: 270–279. doi: 10.1002/jmr.941
|
| [22] |
Ritzefeld M, Walhorn V, Anselmetti D, et al. (2013) Analysis of DNA interactions using single-molecule force spectroscopy. Amino Acids 44: 1457–1475. doi: 10.1007/s00726-013-1474-4
|
| [23] |
Rief M, Clausen-Schaumann H, Gaub HE (1999) Sequence-dependent mechanics of single DNA molecules. Nat Struct Biol 6: 346–349. doi: 10.1038/7582
|
| [24] |
Carrion-Vazquez M, Oberhauser AF, Fowler SB, et al. (1999) Mechanical and chemical unfolding of a single protein: A comparison. Proc Natl Acad Sci U. S. A 96: 3694–3699. doi: 10.1073/pnas.96.7.3694
|
| [25] |
Woodside MT, Block SM (2014) Reconstructing Folding Energy Landscapes by Single-Molecule Force Spectroscopy. Annu Rev Biophys 43: 19–39. doi: 10.1146/annurev-biophys-051013-022754
|
| [26] |
Hughes ML, Dougan L (2016) The physics of pulling polyproteins: A review of single molecule force spectroscopy using the AFM to study protein unfolding. Rep Prog Phys Phys Soc 79: 076601. doi: 10.1088/0034-4885/79/7/076601
|
| [27] |
Fisher TE, Marszalek PE, Fernandez JM (2000) Stretching single molecules into novel conformations using the atomic force microscope. Nat Struct Biol 7: 719–724. doi: 10.1038/78936
|
| [28] |
Beckwitt EC, Kong M, Van Houten B (2018) Studying protein-DNA interactions using atomic force microscopy. Semin Cell Dev Biol 73: 220–230. doi: 10.1016/j.semcdb.2017.06.028
|
| [29] |
Kasas S, Dietler G (2018) DNA-protein interactions explored by atomic force microscopy. Semin Cell Dev Biol 73: 231–239. doi: 10.1016/j.semcdb.2017.07.015
|
| [30] |
Lyubchenko YL, Shlyakhtenko LS (2016) Imaging of DNA and Protein-DNA Complexes with Atomic Force Microscopy. Crit Rev Eukaryot Gene Expr 26: 63–96. doi: 10.1615/CritRevEukaryotGeneExpr.v26.i1.70
|
| [31] |
Halford SE, Marko JF (2004) How do site-specific DNA-binding proteins find their targets? Nucleic Acids Res 32: 3040–3052. doi: 10.1093/nar/gkh624
|
| [32] |
Schneider R, Grosschedl R (2007) Dynamics and interplay of nuclear architecture, genome organization, and gene expression. Genes Dev 21: 3027–3043. doi: 10.1101/gad.1604607
|
| [33] |
Lambert SA, Jolma A, Campitelli LF, et al. (2018) The Human Transcription Factors. Cell 172: 650–665. doi: 10.1016/j.cell.2018.01.029
|
| [34] |
Sancar A, Lindsey-Boltz LA, Unsal-Kacmaz K, et al. (2004) Molecular mechanisms of mammalian DNA repair and the DNA damage checkpoints. Annu Rev Biochem 73: 39–85. doi: 10.1146/annurev.biochem.73.011303.073723
|
| [35] |
Kapanidis AN, Strick T (2009) Biology, one molecule at a time. Trends Biochem Sci 34: 234–243. doi: 10.1016/j.tibs.2009.01.008
|
| [36] |
Larson MH, Landick R, Block SM (2011) Single-Molecule Studies of RNA Polymerase: One Singular Sensation, Every Little Step It Takes. Mol Cell 41: 249–262. doi: 10.1016/j.molcel.2011.01.008
|
| [37] |
Ghodke H, Wang H, Hsieh CL, et al. (2014) Single-molecule analysis reveals human UV-damaged DNA-binding protein (UV-DDB) dimerizes on DNA via multiple kinetic intermediates. P Natl Acad Sci USA 111: E1862–E1871. doi: 10.1073/pnas.1323856111
|
| [38] |
Buechner CN, Maiti A, Drohat AC, et al. (2015) Lesion search and recognition by thymine DNA glycosylase revealed by single molecule imaging. Nucleic Acids Res 43: 2716–2729. doi: 10.1093/nar/gkv139
|
| [39] |
Bewley CA, Gronenborn AM, Clore GM (1998) Minor groove-binding architectural proteins: Structure, function, and DNA recognition. Annu Rev Biophys Biomol Struct 27: 105–131. doi: 10.1146/annurev.biophys.27.1.105
|
| [40] |
Perez-Rueda E, Hernandez-Guerrero R, Martinez-Nunez MA, et al. (2018) Abundance, diversity and domain architecture variability in prokaryotic DNA-binding transcription factors. PLos One 13: e0195332. doi: 10.1371/journal.pone.0195332
|
| [41] |
Richards FM, Kundrot CE (1988) Identification of Structural Motifs from Protein Coordinate Data-Secondary Structure and 1st-Level Supersecondary Structure. Proteins 3: 71–84. doi: 10.1002/prot.340030202
|
| [42] |
Shanahan HP, Garcia MA, Jones S, et al. (2004) Identifying DNA-binding proteins using structural motifs and the electrostatic potential. Nucleic Acids Res 32: 4732–4741. doi: 10.1093/nar/gkh803
|
| [43] |
Bustin M, Reeves R (1996) High-mobility-group chromosomal proteins: Architectural components that facilitate chromatin function. Prog Nucleic Acid Res Mol Biol 54: 35–100. doi: 10.1016/S0079-6603(08)60360-8
|
| [44] |
Smith NC, Matthews JM (2016) Mechanisms of DNA-binding specificity and functional gene regulation by transcription factors. Curr Opin Struct Biol 38: 68–74. doi: 10.1016/j.sbi.2016.05.006
|
| [45] |
Saravanan M, Vasu K, Nagaraja V (2008) Evolution of sequence specificity in a restriction endonuclease by a point mutation. Proc Natl Acad Sci U. S. A 105: 10344–10347. doi: 10.1073/pnas.0804974105
|
| [46] |
Ferredamare AR, Prendergast GC, Ziff EB, et al. (1993) Recognition by Max of Its Cognate DNA through a Dimeric B/Hlh/Z Domain. Nature 363: 38–45. doi: 10.1038/363038a0
|
| [47] |
Morgunova E, Yin Y, Jolma A, et al. (2015) Structural insights into the DNA-binding specificity of E2F family transcription factors. Nat Commun 6: 10050. doi: 10.1038/ncomms10050
|
| [48] |
Li J, Rodriguez JP, Niu F, et al. (2016) Structural basis for DNA recognition by STAT6. P Natl Acad Sci USA 113: 13015–13020. doi: 10.1073/pnas.1611228113
|
| [49] |
Rudolph MJ, Gergen JP (2001) DNA-binding by Ig-fold proteins. Nat Struct Biol 8: 384–386. doi: 10.1038/87531
|
| [50] |
Doherty AJ, Serpell LC, Ponting CP (1996) The helix-hairpin-helix DNA-binding motif: A structural basis for non-sequence-specific recognition of DNA. Nucleic Acids Res 24: 2488–2497. doi: 10.1093/nar/24.13.2488
|
| [51] |
Burgers PMJ, Kunkel TA (2017) Eukaryotic DNA Replication Fork. Annu Rev Biochem 86: 417–438. doi: 10.1146/annurev-biochem-061516-044709
|
| [52] |
Raghunathan S, Kozlov AG, Lohman TM, et al. (2000) Structure of the DNA binding domain of E-coli SSB bound to ssDNA. Nat Struct Biol 7: 648–652. doi: 10.1038/77943
|
| [53] |
Theis K, Chen PJ, Skorvaga M, et al. (1999) Crystal structure of UvrB, a DNA helicase adapted for nucleotide excision repair. EMBO J 18: 6899–6907. doi: 10.1093/emboj/18.24.6899
|
| [54] |
Waters TR, Eryilmaz J, Geddes S, et al. (2006) Damage detection by the UvrABC pathway: Crystal structure of UvrB bound to fluorescein-adducted DNA. FEBS Lett 580: 6423–6427. doi: 10.1016/j.febslet.2006.10.051
|
| [55] |
Chai N, Li WX, Wang J, et al. (2015) Structural basis for the Smad5 MH1 domain to recognize different DNA sequences. Nucleic Acids Res 43: 9051–9064. doi: 10.1093/nar/gkv848
|
| [56] |
Maiti A, Morgan MT, Pozharski E, et al. (2008) Crystal structure of human thymine DNA glycosylase bound to DNA elucidates sequence-specific mismatch recognition. P Natl Acad Sci USA 105: 8890–8895. doi: 10.1073/pnas.0711061105
|
| [57] |
Bruner SD, Norman DPG, Verdine GL (2000) Structural basis for recognition and repair of the endogenous mutagen 8-oxoguanine in DNA. Nature 403: 859–866. doi: 10.1038/35002510
|
| [58] |
Ando T, Kodera N, Takai E, et al. (2001) A high-speed atomic force microscope for studying biological macromolecules. P Natl Acad Sci USA 98: 12468–12472. doi: 10.1073/pnas.211400898
|
| [59] |
Ando T (2018) High-speed atomic force microscopy and its future prospects. Biophys Rev 10: 285–292. doi: 10.1007/s12551-017-0356-5
|
| [60] |
Sawicka M, Aramayo R, Ayala R, et al. (2017) Single-Particle Electron Microscopy Analysis of DNA Repair Complexes. Methods Enzymol 592: 159–186. doi: 10.1016/bs.mie.2017.03.010
|
| [61] |
Sun JC, Yuan ZN, Bai L, et al. (2017) Cryo-EM of dynamic protein complexes in eukaryotic DNA replication. Protein Sci 26: 40–51. doi: 10.1002/pro.3033
|
| [62] |
Tessmer I, Moore T, Lloyd RG, et al. (2005) AFM studies on the role of the protein RdgC in bacterial DNA recombination. J Mol Biol 350: 254–262. doi: 10.1016/j.jmb.2005.04.043
|
| [63] |
Lohr D, Bash R, Wang H, et al. (2007) Using atomic force microscopy to study chromatin structure and nucleosome remodeling. Methods 41: 333–341. doi: 10.1016/j.ymeth.2006.08.016
|
| [64] |
Hizume K, Kominam H, Kobayash K, et al. (2017) Flexible DNA Path in the MCM Double Hexamer Loaded on DNA. Biochemistry 56: 2435–2445. doi: 10.1021/acs.biochem.6b00922
|
| [65] |
Noort JV, Heijden TVD, Dutta CF, et al. (2004) Initiation of translocation by Type I restriction-modification enzymes is associated with a short DNA extrusion. Nucleic Acids Res 32: 6540–6547. doi: 10.1093/nar/gkh999
|
| [66] |
Maurer S, Fritz J, Muskhelishvili G, et al. (2006) RNA polymerase and an activator form discrete subcomplexes in a transcription initiation complex. EMBO J 25: 3784–3790. doi: 10.1038/sj.emboj.7601261
|
| [67] |
Verhoeven EEA, Wyman C, Moolenaar GF, et al. (2002) The presence of two UvrB subunits in the UvrAB complex ensures damage detection in both DNA strands. EMBO J 21: 4196–4205. doi: 10.1093/emboj/cdf396
|
| [68] |
Cellai S, Mangiarotti L, Vannini N, et al. (2007) Upstream promoter sequences and alpha CTD mediate stable DNA wrapping within the RNA polymerase-promoter open complex. EMBO Rep 8: 271–278. doi: 10.1038/sj.embor.7400888
|
| [69] | Umemura K, Okada T, Kuroda R (2005) Cooperativity and intermediate structures of single-stranded DNA binding-assisted RecA-single-stranded DNA complex formation studied by atomic force microscopy. Scanning 27: 35–43. |
| [70] |
Hamon L, Pastre D, Dupaigne P, et al. (2007) High-resolution AFM imaging of single-stranded DNA-binding (SSB) protein-DNA complexes. Nucleic Acids Res 35: e58. doi: 10.1093/nar/gkm147
|
| [71] |
Li BS, Goh MC (2010) Direct visualization of the formation and structure of RecA/dsDNA complexes. Micron 41: 227–231. doi: 10.1016/j.micron.2009.10.011
|
| [72] |
Li BS, Wei B, Goh MC (2012) Direct visualization of the formation of RecA/dsDNA complexes at the single-molecule level. Micron 43: 1073–1075. doi: 10.1016/j.micron.2012.04.016
|
| [73] |
Tessmer I, Melikishvili M, Fried MG (2012) Cooperative cluster formation, DNA bending and base-flipping by O6-alkylguanine-DNA alkyltransferase. Nucleic Acids Res 40: 8296–8308. doi: 10.1093/nar/gks574
|
| [74] |
Miyagi A, Ando T, Lyubchenko YL (2011) Dynamics of Nucleosomes Assessed with Time-Lapse High-Speed Atomic Force Microscopy. Biochemistry 50: 7901–7908. doi: 10.1021/bi200946z
|
| [75] |
Sanchez H, Suzuki Y, Yokokawa M, et al. (2011) Protein-DNA interactions in high speed AFM: Single molecule diffusion analysis of human RAD54. Integr Biol 3: 1127–1134. doi: 10.1039/c1ib00039j
|
| [76] |
Endo M, Sugiyama H (2014) Single-Molecule Imaging of Dynamic Motions of Biomolecules in DNA Origami Nanostructures Using High-Speed Atomic Force Microscopy. Acc Chem Res 47: 1645–1653. doi: 10.1021/ar400299m
|
| [77] |
Lee AJ, Endo M, Hobbs JK, et al. (2018) Direct Single-Molecule Observation of Mode and Geometry of RecA-Mediated Homology Search. Acs Nano 12: 272–278. doi: 10.1021/acsnano.7b06208
|
| [78] |
Suzuki Y, Shin M, Yoshida A, et al. (2012) Fast microscopical dissection of action scenes played by Escherichia coli RNA polymerase. FEBS Lett 586: 3187–3192. doi: 10.1016/j.febslet.2012.06.033
|
| [79] |
Buechner CN, Tessmer I (2013) DNA substrate preparation for atomic force microscopy studies of protein-DNA interactions. J Mol Recognit 26: 605–617. doi: 10.1002/jmr.2311
|
| [80] |
Yang Y, Sass LE, Du C, et al. (2005) Determination of protein-DNA binding constants and specificities from statistical analyses of single molecules: MutS-DNA interactions. Nucleic Acids Res 33: 4322–4334. doi: 10.1093/nar/gki708
|
| [81] |
Sukhanova MV, Abrakhi S, Joshi V, et al. (2016) Single molecule detection of PARP1 and PARP2 interaction with DNA strand breaks and their poly(ADP-ribosyl)ation using high-resolution AFM imaging. Nucleic Acids Res 44: e60. doi: 10.1093/nar/gkv1476
|
| [82] |
Buechner CN, Heil K, Michels G, et al. (2014) Strand-specific Recognition of DNA Damages by XPD Provides Insights into Nucleotide Excision Repair Substrate Versatility. J Biol Chem 289: 3613–3624. doi: 10.1074/jbc.M113.523001
|
| [83] |
Wirth N, Gross J, Roth HM, et al. (2016) Conservation and Divergence in Nucleotide Excision Repair Lesion Recognition. J Biol Chem 291: 18932–18946. doi: 10.1074/jbc.M116.739425
|
| [84] |
Doniselli N, Rodriguez-Aliaga P, Amidani D, et al. (2015) New insights into the regulatory mechanisms of ppGpp and DksA on Escherichia coli RNA polymerase-promoter complex. Nucleic Acids Res 43: 5249–5262. doi: 10.1093/nar/gkv391
|
| [85] |
Nettikadan S, Tokumasu F, Takeyasu K (1996) Quantitative analysis of the transcription factor AP2 binding to DNA by atomic force microscopy. Biochem Biophys Res Commun 226: 645–649. doi: 10.1006/bbrc.1996.1409
|
| [86] |
Timofeeva OA, Chasovskikh S, Lonskaya I, et al. (2012) Mechanisms of Unphosphorylated STAT3 Transcription Factor Binding to DNA. J Biol Chem 287: 14192–14200. doi: 10.1074/jbc.M111.323899
|
| [87] |
Zhang P, Xia JH, Zhu J, et al. (2018) High-throughput screening of prostate cancer risk loci by single nucleotide polymorphisms sequencing. Nat Commun 9: 2022. doi: 10.1038/s41467-018-04451-x
|
| [88] |
Huang Q, Whitington T, Gao P, et al. (2014) A prostate cancer susceptibility allele at 6q22 increases RFX6 expression by modulating HOXB13 chromatin binding. Nat Genet 46: 126–135. doi: 10.1038/ng.2862
|
| [89] |
Gao P, Xia JH, Sipeky C, et al. (2018) Biology and Clinical Implications of the 19q13 Aggressive Prostate Cancer Susceptibility Locus. Cell 174: 576–589. doi: 10.1016/j.cell.2018.06.003
|
| [90] |
Crampton N, Bonass WA, Kirkham J, et al. (2006) Collision events between RNA polymerases in convergent transcription studied by atomic force microscopy. Nucleic Acids Res 34: 5416–5425. doi: 10.1093/nar/gkl668
|
| [91] |
Countryman P, Fan Y, Gorthi A, et al. (2018) Cohesin SA2 is a sequence-independent DNA-binding protein that recognizes DNA replication and repair intermediates. J Biol Chem 293: 1054–1069. doi: 10.1074/jbc.M117.806406
|
| [92] |
Schneider SW, Larmer J, Henderson RM, et al. (1998) Molecular weights of individual proteins correlate with molecular volumes measured by atomic force microscopy. Pflug Arch Eur J Phy 435: 362–367. doi: 10.1007/s004240050524
|
| [93] |
Ratcliff GC, Erie DA (2001) A novel single-molecule study to determine protein-protein association constants. J Am Chem Soc 123: 5632–5635. doi: 10.1021/ja005750n
|
| [94] |
Wang H, Dellavecchia MJ, Skorvaga M, et al. (2006) UvrB domain 4, an autoinhibitory gate for regulation of DNA binding and ATPase activity. J Biol Chem 281: 15227–15237. doi: 10.1074/jbc.M601476200
|
| [95] |
Roth HM, Romer J, Grundler V, et al. (2012) XPB helicase regulates DNA incision by the Thermoplasma acidophilum endonuclease Bax1. DNA Repair 11: 286–293. doi: 10.1016/j.dnarep.2011.12.002
|
| [96] |
Fuentes-Perez ME, Dillingham MS, Moreno-Herrero F (2013) AFM volumetric methods for the characterization of proteins and nucleic acids. Methods 60: 113–121. doi: 10.1016/j.ymeth.2013.02.005
|
| [97] | Amidani D, Tramonti A, Canosa AV, et al. (2016) Study of DNA binding and bending by Bacillus subtilis GabR, a PLP-dependent transcription factor. Biochim Biophys Acta Gen Subj 1861: 3474–3489. |
| [98] |
Rivetti C, Guthold M, Bustamante C (1996) Scanning force microscopy of DNA deposited onto mica: Equilibration versus kinetic trapping studied by statistical polymer chain analysis. J Mol Biol 264: 919–932. doi: 10.1006/jmbi.1996.0687
|
| [99] |
Cassina V, Manghi M, Salerno D, et al. (2016) Effects of cytosine methylation on DNA morphology: An atomic force microscopy study. Biochim Biophys Acta 1860: 1–7. doi: 10.1016/j.bbagen.2015.10.006
|
| [100] |
Scipioni A, Anselmi C, Zuccheri G, et al. (2002) Sequence-dependent DNA curvature and flexibility from scanning force microscopy images. Biophys J 83: 2408–2418. doi: 10.1016/S0006-3495(02)75254-5
|
| [101] |
Moukhtar J, Faivre-Moskalenko C, Milani P, et al. (2010) Effect of genomic long-range correlations on DNA persistence length: from theory to single molecule experiments. J Phys Chem B 114: 5125–5143. doi: 10.1021/jp911031y
|
| [102] |
Jager MD, Noort JV, Gent DCV, et al. (2001) Human Rad50/Mre11 is a flexible complex that can tether DNA ends. Mol Cell 8: 1129–1135. doi: 10.1016/S1097-2765(01)00381-1
|
| [103] |
Tessmer I, Yang Y, Zhai J, et al. (2008) Mechanism of MutS searching for DNA mismatches and signaling repair. J Biol Chem 283: 36646–36654. doi: 10.1074/jbc.M805712200
|
| [104] |
Bosch D, Campillo M, Pardo L (2003) Binding of proteins to the minor groove of DNA: What are the structural and energetic determinants for kinking a basepair step? J Comput Chem 24: 682–691. doi: 10.1002/jcc.10200
|
| [105] |
Kong MW, Liu LL, Chen XJ, et al. (2016) Single-Molecule Imaging Reveals that Rad4 Employs a Dynamic DNA Damage Recognition Process. Mol Cell 64: 376–387. doi: 10.1016/j.molcel.2016.09.005
|
| [106] |
Wang H, Yang Y, Schofield MJ, et al. (2003) DNA bending and unbending by MutS govern mismatch recognition and specificity. Proc Natl Acad Sci U. S. A 100: 14822–14827. doi: 10.1073/pnas.2433654100
|
| [107] |
Chen L, Haushalter KA, Lieber CM, et al. (2002) Direct visualization of a DNA glycosylase searching for damage. Chem Biol 9: 345–350. doi: 10.1016/S1074-5521(02)00120-5
|
| [108] |
Lamers MH, Perrakis A, Enzlin JH, et al. (2000) The crystal structure of DNA mismatch repair protein MutS binding to a G center dot T mismatch. Nature 407: 711–717. doi: 10.1038/35037523
|
| [109] |
Koroleva ON, Dubrovin EV, Yaminsky IV, et al. (2016) Effect of DNA bending on transcriptional interference in the systems of closely spaced convergent promoters. Biochim Biophys Acta 1860: 2086–2096. doi: 10.1016/j.bbagen.2016.06.026
|
| [110] |
Fronczek DN, Quammen C, Wang H, et al. (2011) High accuracy FIONA-AFM hybrid imaging. Ultramicroscopy 111: 350–355. doi: 10.1016/j.ultramic.2011.01.020
|
| [111] |
Sanchez H, Kertokalio A, van Rossum-Fikkert S, et al. (2013) Combined optical and topographic imaging reveals different arrangements of human RAD54 with presynaptic and postsynaptic RAD51-DNA filaments. P Natl Acad Sci USA 110: 11385–11390. doi: 10.1073/pnas.1306467110
|
| [112] |
Frederickx W, Rocha S, Fujita Y, et al. (2018) Orthogonal Probing of Single-Molecule Heterogeneity by Correlative Fluorescence and Force Microscopy. Acs Nano 12: 168–177. doi: 10.1021/acsnano.7b05405
|
| [113] |
Schmucker SW, Kumar N, Abelson JR, et al. (2012) Field-directed sputter sharpening for tailored probe materials and atomic-scale lithography. Nat Commun 3: 935. doi: 10.1038/ncomms1907
|
| [114] |
Pfreundschuh M, Alsteens D, Hilbert M, et al. (2014) Localizing Chemical Groups while Imaging Single Native Proteins by High-Resolution Atomic Force Microscopy. Nano Lett 14: 2957–2964. doi: 10.1021/nl5012905
|
| [115] |
Monig H, Hermoso DR, Arado OD, et al. (2016) Submolecular Imaging by Noncontact Atomic Force Microscopy with an Oxygen Atom Rigidly Connected to a Metallic Probe. Acs Nano 10: 1201–1209. doi: 10.1021/acsnano.5b06513
|
| [116] |
Senapati S, Lindsay S (2016) Recent Progress in Molecular Recognition Imaging Using Atomic Force Microscopy. Acc Chem Res 49: 503–510. doi: 10.1021/acs.accounts.5b00533
|
| [117] | Khan Z, Leung C, Tahir BA, et al. (2010) Digitally tunable, wide-band amplitude, phase, and frequency detection for atomic-resolution scanning force microscopy. Rev Sci Instrum 81: 197. |
| [118] |
Calo A, Eleta-Lopez A, Stoliar P, et al. (2016) Multifrequency Force Microscopy of Helical Protein Assembly on a Virus. Sci Rep 6: 21899. doi: 10.1038/srep21899
|
| [119] |
Wu D, Kaur P, Li ZM, et al. (2016) Visualizing the Path of DNA through Proteins Using DREEM Imaging. Mol Cell 61: 315–323. doi: 10.1016/j.molcel.2015.12.012
|
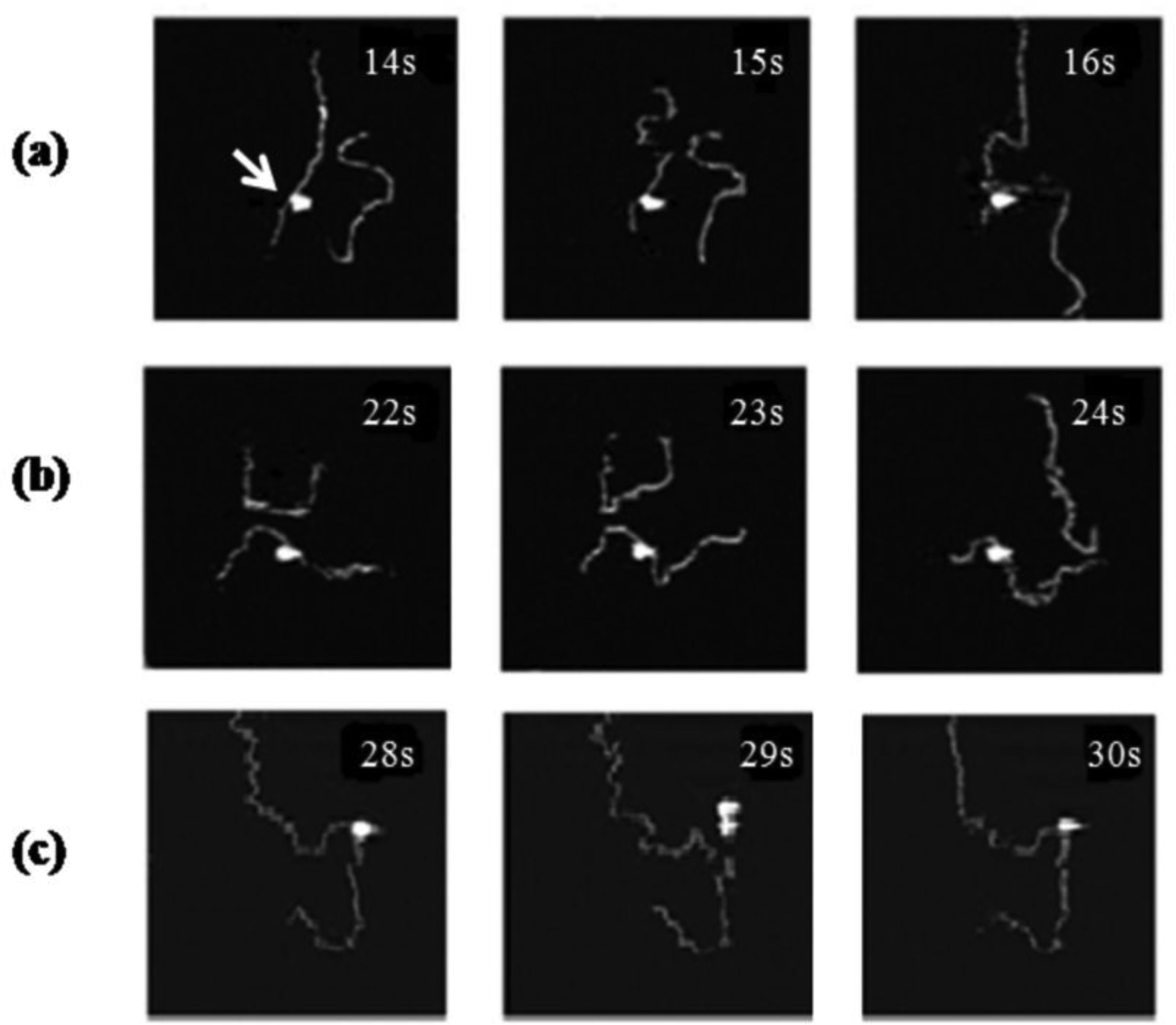
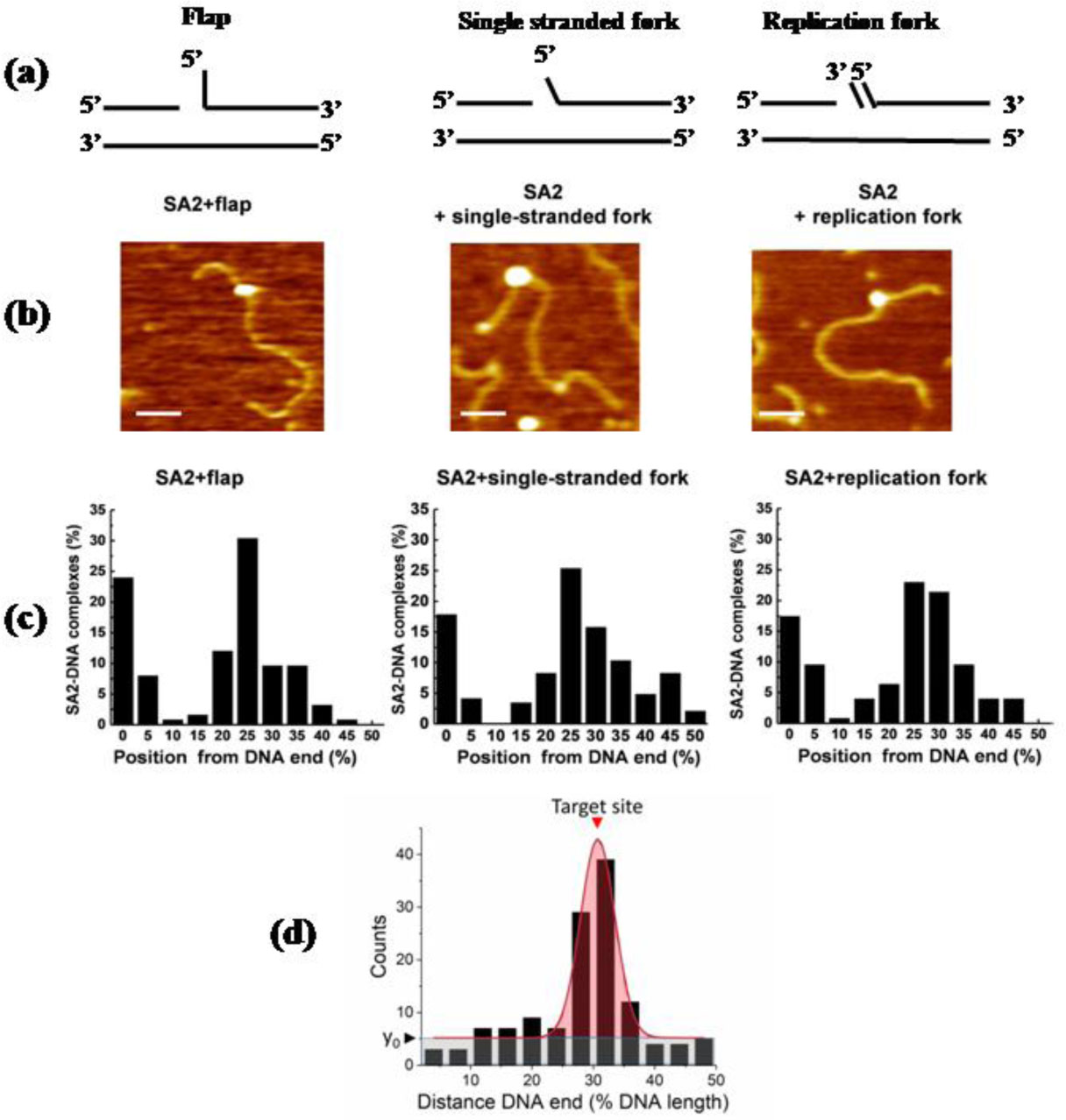
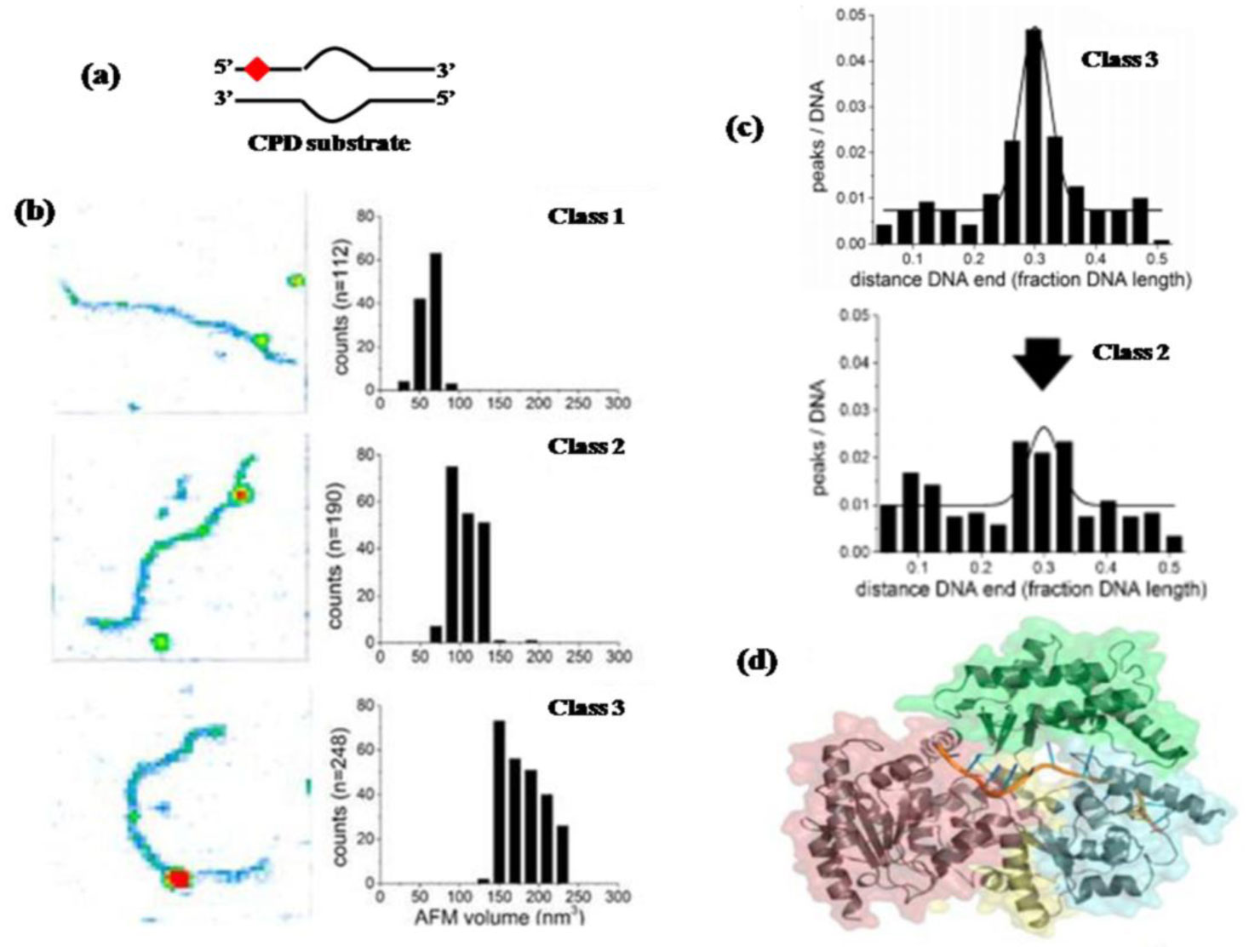
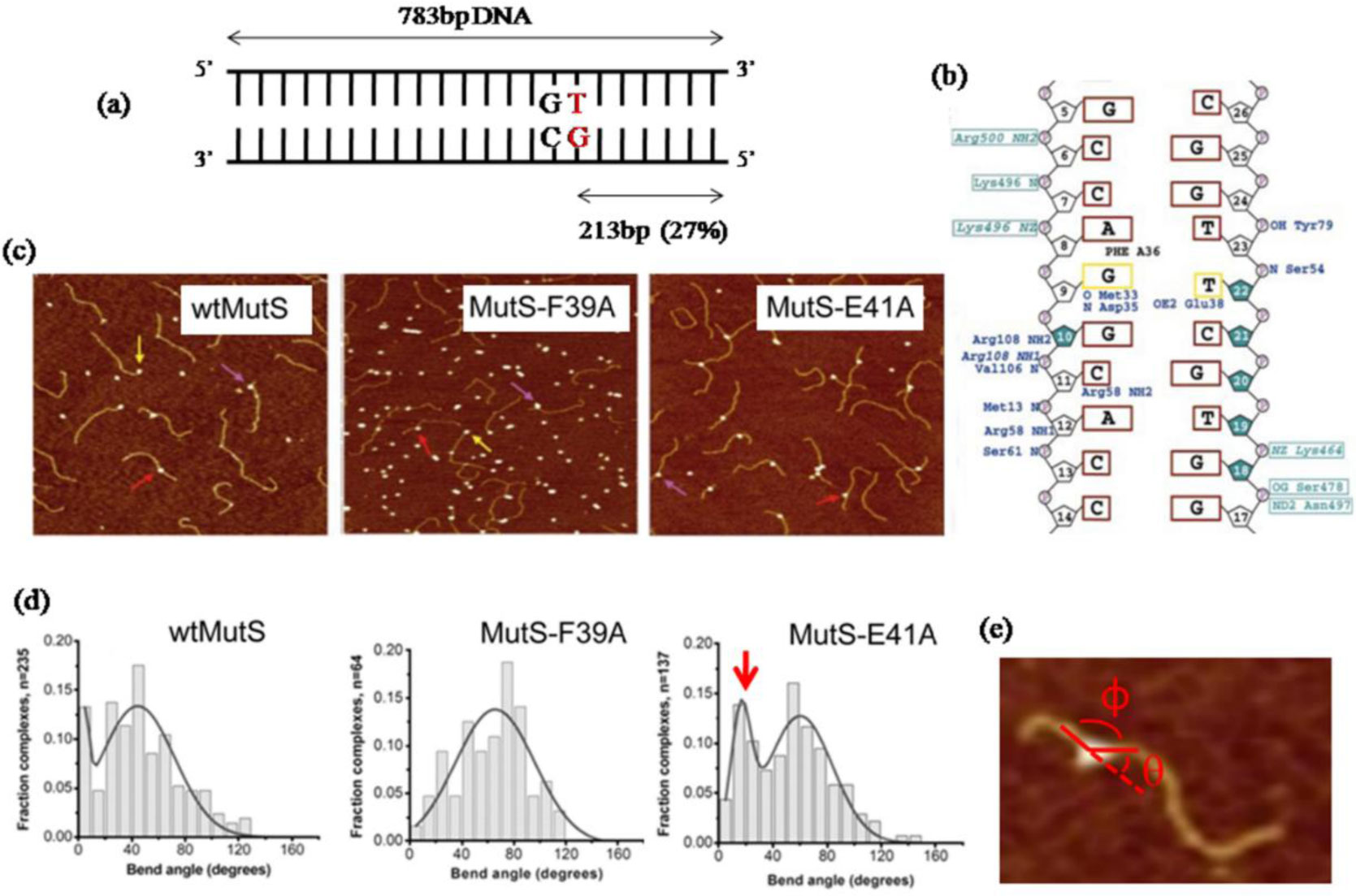
Figures(8)

Disha Mohan Bangalore, Ingrid Tessmer. Unique insight into protein-DNA interactions from single molecule atomic force microscopy[J]. AIMS Biophysics, 2018, 5(3): 194-216. doi: 10.3934/biophy.2018.3.194







 DownLoad:
DownLoad: