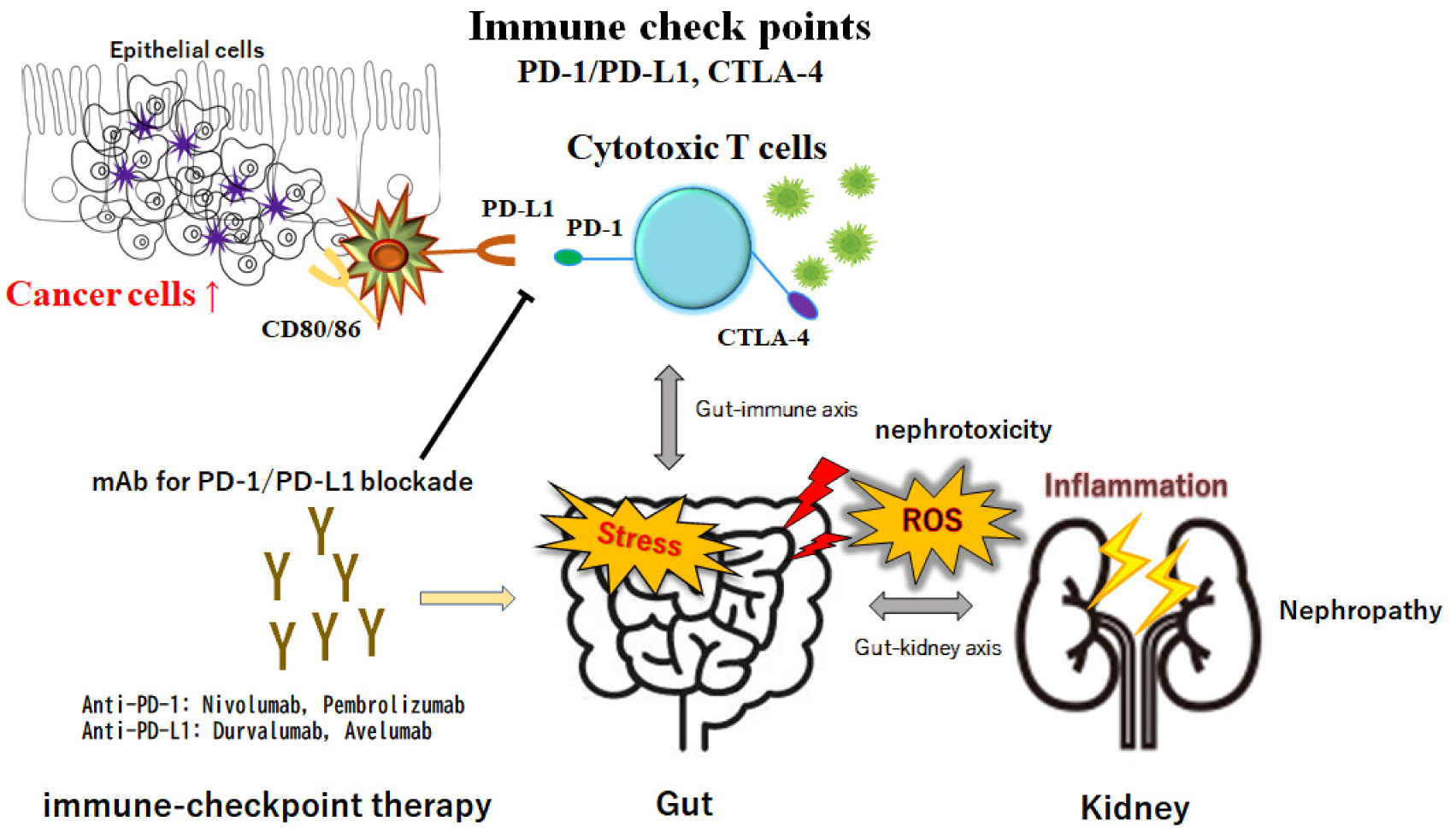
The immune-related adverse events resulting from the therapy of immune checkpoint inhibitors could cause kidney injury. Inflammatory reprogramming of regulatory T helper (Treg) cells or type 17 T helper (Th17) cells might be involved in the pathogenesis of nephropathy. Accumulating evidence confirms a connection between the diversity of gut microbiota and kidney diseases, suggesting that successful modification of gut microbiota could attenuate kidney injury. In other words, certain gut microbiota could contribute to the protection of kidneys via the gut-kidney axis. It has been shown that the dysbiosis of gut microbiota might affect the gut-kidney axis, leading to nephrotoxicity. On the contrary, altered levels of D-amino acids, ROS, and SCFAs through the adjustment of gut microbiota might be relevant to the reduction of nephrotoxicity. Here, we have discussed and suggested the beneficial roles of gut microbiota in the prevention of the kidney injury induced during immune-checkpoint therapy.
Citation: Sayuri Yoshikawa, Kurumi Taniguchi, Haruka Sawamura, Yuka Ikeda, Ai Tsuji, Satoru Matsuda. Promising probiotics for the treatment of nephrotoxicity induced during immune-checkpoint therapy against cancers[J]. AIMS Bioengineering, 2022, 9(3): 283-292. doi: 10.3934/bioeng.2022019
The immune-related adverse events resulting from the therapy of immune checkpoint inhibitors could cause kidney injury. Inflammatory reprogramming of regulatory T helper (Treg) cells or type 17 T helper (Th17) cells might be involved in the pathogenesis of nephropathy. Accumulating evidence confirms a connection between the diversity of gut microbiota and kidney diseases, suggesting that successful modification of gut microbiota could attenuate kidney injury. In other words, certain gut microbiota could contribute to the protection of kidneys via the gut-kidney axis. It has been shown that the dysbiosis of gut microbiota might affect the gut-kidney axis, leading to nephrotoxicity. On the contrary, altered levels of D-amino acids, ROS, and SCFAs through the adjustment of gut microbiota might be relevant to the reduction of nephrotoxicity. Here, we have discussed and suggested the beneficial roles of gut microbiota in the prevention of the kidney injury induced during immune-checkpoint therapy.
adenosine triphosphate
cytotoxic T lymphocyte-associated protein 4
chronic kidney disease
deoxyribonucleic acid
fecal microbiota transplantation
graft-versus-host disease
histone deacetylase
programmed cell death protein 1
programmed cell death ligand 1
reactive oxygen species
superoxide dismutase
type 1 T helper
type 17 T helper
regulatory T

| [1] |
Min JW, Lim JU (2022) Immune checkpoint inhibitors in patients with chronic kidney disease: Assessing their ability to cause acute kidney injury and informing their proper use. Semin Oncol 49: 141-147. https://doi.org/10.1053/j.seminoncol.2022.01.012 
|
| [2] |
Seethapathy H, Herrmann SM, Sise ME (2021) Immune checkpoint inhibitors and kidney toxicity: advances in diagnosis and management. Kidney Med 3: 1074-1081. https://doi.org/10.1016/j.xkme.2021.08.008
|
| [3] |
Di Giacomo AM, Guarnieri A, Tripodi SA, et al. (2022) Brief communication PD1-related nephrotoxicity: optimizing its clinical management through histopathologic features. J Immunother 45: 217-221. https://doi.org/10.1097/CJI.0000000000000412
|
| [4] |
Chang CY, Park H, Malone DC, et al. (2020) Immune checkpoint inhibitors and immune-related adverse events in patients with advanced melanoma: a systematic review and network meta-analysis. JAMA Netw Open 3: e201611. https://doi.org/10.1001/jamanetworkopen.2020.1611
|
| [5] |
Yang J, Ji GE, Park MS, et al. (2021) Probiotics partially attenuate the severity of acute kidney injury through an immunomodulatory effect. Kidney Res Clin Pract 40: 620-633. https://doi.org/10.23876/j.krcp.20.265
|
| [6] |
Lin D, Hu B, Li P, et al. (2021) Roles of the intestinal microbiota and microbial metabolites in acute GVHD. Exp Hematol Oncol 10: 49. https://doi.org/10.1186/s40164-021-00240-3
|
| [7] |
Fan Z, Ross RP, Stanton C, et al. (2021) Lactobacillus casei CCFM1074 alleviates collagen-induced arthritis in rats via balancing Treg/Th17 and modulating the metabolites and gut microbiota. Front Immunol 12: 680073. https://doi.org/10.3389/fimmu.2021.680073
|
| [8] |
Sumida K, Lau WL, Kovesdy CP, et al. (2021) Microbiome modulation as a novel therapeutic approach in chronic kidney disease. Curr Opin Nephrol Hypertens 30: 75-84. https://doi.org/10.1097/MNH.0000000000000661
|
| [9] |
Lapaquette P, Bizeau JB, Acar N, et al. (2021) Reciprocal interactions between gut microbiota and autophagy. World J Gastroenterol 27: 8283-8301. https://doi.org/10.3748/wjg.v27.i48.8283
|
| [10] |
Gambichler T, Schröter U, Höxtermann S, et al. (2020) Decline of programmed death-1-positive circulating T regulatory cells predicts more favourable clinical outcome of patients with melanoma under immune checkpoint blockade. Br J Dermatol 182: 1214-1220. https://doi.org/10.1111/bjd.18379
|
| [11] |
Sandin LC, Eriksson F, Ellmark P, et al. (2014) Local CTLA4 blockade effectively restrains experimental pancreatic adenocarcinoma growth in vivo. Oncoimmunology 3: e27614. https://doi.org/10.4161/onci.27614
|
| [12] |
Kumar P, Saini S, Prabhakar BS (2020) Cancer immunotherapy with check point inhibitor can cause autoimmune adverse events due to loss of Treg homeostasis. Semin Cancer Biol 64: 29-35. https://doi.org/10.1016/j.semcancer.2019.01.006
|
| [13] |
Okiyama N, Tanaka R (2022) Immune-related adverse events in various organs caused by immune checkpoint inhibitors. Allergol Int 71: 169-178. https://doi.org/10.1016/j.alit.2022.01.001
|
| [14] |
Sun S, Wang F (2020) Molecular events behind adverse effects. Adv Exp Med Biol 1248: 119-141. https://doi.org/10.1007/978-981-15-3266-5_6
|
| [15] |
Maillard A, Pastor D, Merat R (2021) Anti-PD-1-Induced hidradenitis suppurativa. Dermatopathology 8: 37-39. https://doi.org/10.3390/dermatopathology8010007
|
| [16] |
Gao Y, Shan W, Gu T, et al. (2021) Daratumumab prevents experimental xenogeneic graft-versus-host disease by skewing proportions of T Cell functional subsets and inhibiting T Cell activation and migration. Front Immunol 12: 785774. https://doi.org/10.3389/fimmu.2021.785774
|
| [17] |
Hailemichael Y, Johnson DH, Abdel-Wahab N, et al. (2022) Interleukin-6 blockade abrogates immunotherapy toxicity and promotes tumor immunity. Cancer Cell 40: 509-523.E6. https://doi.org/10.1016/j.ccell.2022.04.004
|
| [18] |
Kim ST, Chu Y, Misoi M, et al. (2022) Distinct molecular and immune hallmarks of inflammatory arthritis induced by immune checkpoint inhibitors for cancer therapy. Nat Commun 13: 1970. https://doi.org/10.1038/s41467-022-29539-3
|
| [19] |
Li Y, Tang D, Yin L, et al. (2022) New insights for regulatory T cell in lupus nephritis. Autoimmun Rev 21: 103134. https://doi.org/10.1016/j.autrev.2022.103134
|
| [20] |
Wang B, Jiang X, Li Y, et al. (2022) YY1 alleviates lupus nephritis-induced renal injury by reducing the Th17/Treg cell ratio via the IFN-γ/Fra2 axis. Lab Invest 102: 872-884. https://doi.org/10.1038/s41374-022-00777-9
|
| [21] |
Turner JE, Paust HJ, Steinmetz OM, et al. (2010) The Th17 immune response in renal inflammation. Kidney Int 77: 1070-1075. https://doi.org/10.1038/ki.2010.102
|
| [22] |
Duffy MM, McNicholas BA, Monaghan DA, et al. (2014) Mesenchymal stem cells and a vitamin D receptor agonist additively suppress T helper 17 cells and the related inflammatory response in the kidney. Am J Physiol Renal Physiol 307: F1412-F1426. https://doi.org/10.1152/ajprenal.00024.2014
|
| [23] | Li YY, Wei SG, Zhao X, et al. (2016) Th17/Treg cell expression in children with primary nephritic syndrome and the effects of ox-LDL on Th17/Treg cells. Genet Mol Res 15: gmr.15027669. https://doi.org/10.4238/gmr.15027669 |
| [24] |
Grigoriou M, Banos A, Hatzioannou A, et al. (2021) Regulatory T-cell transcriptomic reprogramming characterizes adverse events by checkpoint inhibitors in solid tumors. Cancer Immunol Res 9: 726-734. https://doi.org/10.1158/2326-6066.CIR-20-0969
|
| [25] |
Kitai T, Tang WHW (2018) Gut microbiota in cardiovascular disease and heart failure. Clin Sci (Lond) 132: 85-91. https://doi.org/10.1042/CS20171090
|
| [26] |
Sata Y, Marques FZ, Kaye DM (2020) The emerging role of gut dysbiosis in cardio-metabolic risk factors for heart failure. Curr Hypertens Rep 22: 38. https://doi.org/10.1007/s11906-020-01046-0
|
| [27] |
Ballard JWO, Towarnicki SG (2020) Mitochondria, the gut microbiome and ROS. Cell Signal 75: 109737. https://doi.org/10.1016/j.cellsig.2020.109737
|
| [28] |
Espinosa A, Henríquez-Olguín C, Jaimovich E (2016) Reactive oxygen species and calcium signals in skeletal muscle: A crosstalk involved in both normal signaling and disease. Cell Calcium 60: 172-179. https://doi.org/10.1016/j.ceca.2016.02.010
|
| [29] |
Gao L, Luo D, Wu D, et al. (2022) Effects of mammalian target of rapamycin and aryl hydrocarbon receptor-mediating autophagy signaling on the balance of Th17/Treg cells during perinatal bisphenol a exposure in female offspring mice. Environ Toxicol 37: 1781-1789. https://doi.org/10.1002/tox.23525
|
| [30] |
Fernández-Sánchez A, Madrigal-Santillán E, Bautista M, et al. (2011) Inflammation, oxidative stress, and obesity. Int J Mol Sci 12: 3117-3132. https://doi.org/10.3390/ijms12053117
|
| [31] |
Wada A, Higashiyama M, Kurihara C, et al. (2022) Protective effect of luminal uric acid against indomethacin-induced enteropathy: role of antioxidant effect and gut microbiota. Dig Dis Sci 67: 121-133. https://doi.org/10.1007/s10620-021-06848-z
|
| [32] |
Yang T, Richards EM, Pepine CJ, et al. (2018) The gut microbiota and the brain-gut-kidney axis in hypertension and chronic kidney disease. Nat Rev Nephrol 14: 442-456. https://doi.org/10.1038/s41581-018-0018-2
|
| [33] |
Peng SN, Zeng HH, Fu AX, et al. (2013) Effects of rhein on intestinal epithelial tight junction in IgA nephropathy. World J Gastroenterol 19: 4137-4145. https://doi.org/10.3748/wjg.v19.i26.4137
|
| [34] |
Sol CM, Santos S, Asimakopoulos AG, et al. (2020) Associations of maternal phthalate and bisphenol urine concentrations during pregnancy with childhood blood pressure in a population-based prospective cohort study. Environ Int 138: 105677. https://doi.org/10.1016/j.envint.2020.105677
|
| [35] |
Andersen K, Kesper MS, Marschner JA, et al. (2017) Intestinal dysbiosis, barrier dysfunction, and bacterial translocation account for CKD-related systemic inflammation. J Am Soc Nephrol 28: 76-83. https://doi.org/10.1681/ASN.2015111285
|
| [36] |
Nagase N, Ikeda Y, Tsuji A, et al. (2022) Efficacy of probiotics on the modulation of gut microbiota in the treatment of diabetic nephropathy. World J Diabetes 13: 150-160. https://doi.org/10.4239/wjd.v13.i3.150
|
| [37] |
Zhao J, Ning X, Liu B, et al. (2021) Specific alterations in gut microbiota in patients with chronic kidney disease: an updated systematic review. Ren Fail 43: 102-112. https://doi.org/10.1080/0886022X.2020.1864404
|
| [38] |
Han C, Jiang YH, Li W, et al. (2021) Astragalus membranaceus and Salvia miltiorrhiza ameliorates cyclosporin a-induced chronic nephrotoxicity through the “gut-kidney axis”. J Ethnopharmacol 269: 113768. https://doi.org/10.1016/j.jep.2020.113768
|
| [39] |
Bian J, Liebert A, Bicknell B, et al. (2022) Faecal microbiota transplantation and chronic kidney disease. Nutrients 14: 2528. https://doi.org/10.3390/nu14122528
|
| [40] |
Feng Y, Li L, Guo F, et al. (2018) Protective effects of SKLB023 on a mouse model of unilateral ureteral obstruction by the modulation of gut microbiota. RSC Adv 8: 40232-40242. https://doi.org/10.1039/C8RA08049F
|
| [41] |
Campaniello D, Corbo MR, Sinigaglia M, et al. (2022) How diet and physical activity modulate gut microbiota: evidence and perspectives. Nutrients 14: 2456. https://doi.org/10.3390/nu14122456
|
| [42] |
Zheng HJ, Guo J, Wang Q, et al. (2021) Probiotics, prebiotics, and synbiotics for the improvement of metabolic profiles in patients with chronic kidney disease: A systematic review and meta-analysis of randomized controlled trials. Crit Rev Food Sci Nutr 61: 577-598. https://doi.org/10.1080/10408398.2020.1740645
|
| [43] |
Pandey KR, Naik SR, Vakil BV (2015) Probiotics, prebiotics and synbiotics- a review. J Food Sci Technol 52: 7577-7587. https://doi.org/10.1007/s13197-015-1921-1
|
| [44] |
Żółkiewicz J, Marzec A, Ruszczyński M, et al. (2020) Postbiotics-a step beyond pre- and probiotics. Nutrients 12: 2189. https://doi.org/10.3390/nu12082189
|
| [45] |
Iwata Y, Nakade Y, Kitajima S, et al. (2022) Protective effect of d-alanine against acute kidney injury. Am J Physiol Renal Physiol 322: F667-F679. https://doi.org/10.1152/ajprenal.00198.2021
|
| [46] |
Nakade Y, Iwata Y, Furuichi K, et al. (2018) Gut microbiota-derived D-serine protects against acute kidney injury. JCI Insight 3: e97957. https://doi.org/10.1172/jci.insight.97957
|
| [47] |
Winiarska-Mieczan A, Tomaszewska E, Donaldson J, et al. (2022) The Role of nutritional factors in the modulation of the composition of the gut microbiota in people with autoimmune diabetes. Nutrients 14: 2498. https://doi.org/10.3390/nu14122498
|
| [48] |
Flores MV, Crawford KC, Pullin LM, et al. (2010) Dual oxidase in the intestinal epithelium of zebrafish larvae has anti-bacterial properties. Biochem Biophys Res Commun 400: 164-168. https://doi.org/10.1016/j.bbrc.2010.08.037
|
| [49] |
Wang T, Yang J, Lin G, et al. (2021) Effects of dietary mannan oligosaccharides on non-specific immunity, intestinal health, and antibiotic resistance genes in pacific white shrimp litopenaeus vannamei. Front Immunol 12: 772570. https://doi.org/10.3389/fimmu.2021.772570
|
| [50] |
Abdel-Fattah MM, Elgendy ANAM, Mohamed WR (2021) X anthenone, ACE2 activator, counteracted gentamicin-induced nephrotoxicity in rats: Impact on oxidative stress and ACE2/Ang-(1-7) signaling. Life Sci 275: 119387. https://doi.org/10.1016/j.lfs.2021.119387
|
| [51] | Ratajczak W, Rył A, Mizerski A, et al. (2019) Immunomodulatory potential of gut microbiome-derived short-chain fatty acids (SCFAs). Acta Biochim Pol 66: 1-12. https://doi.org/10.18388/abp.2018_2648 |
| [52] |
Zhang H, Zhang W, Jiao F, et al. (2018) The nephroprotective effect of MS-275 on lipopolysaccharide (LPS)-induced acute kidney injury by inhibiting reactive oxygen species (ROS)-oxidative stress and endoplasmic reticulum stress. Med Sci Monit 24: 2620-2630. https://doi.org/10.12659/MSM.906362
|
| [53] |
D'Souza WN, Douangpanya J, Mu S, et al. (2017) Differing roles for short chain fatty acids and GPR43 agonism in the regulation of intestinal barrier function and immune responses. PLoS One 12: e0180190. https://doi.org/10.1371/journal.pone.0180190
|
| [54] |
Andrade-Oliveira V, Amano MT, Correa-Costa M, et al. (2015) Gut bacteria products prevent AKI induced by ischemia-reperfusion. J Am Soc Nephrol 26: 1877-1888. https://doi.org/10.1681/ASN.2014030288
|
| [55] |
Sun S, Luo L, Liang W, et al. (2020) Bifidobacterium alters the gut microbiota and modulates the functional metabolism of T regulatory cells in the context of immune checkpoint blockade. Proc Natl Acad Sci U S A 117: 27509-27515. https://doi.org/10.1073/pnas.1921223117
|
| [56] |
Zou YT, Zhou J, Zhu JH, et al. (2022) Gut microbiota mediates the protective effects of traditional chinese medicine formula Qiong-Yu-Gao against cisplatin-induced acute kidney injury. Microbiol Spectr 10: e0075922. https://doi.org/10.1128/spectrum.00759-22
|
| [57] |
Wilck N, Matus MG, Kearney SM, et al. (2017) Salt-responsive gut commensal modulates TH17 axis and disease. Nature 551: 585-589. https://doi.org/10.1038/nature24628
|
| [58] |
Jung SM, Kim Y, Kim J, et al. (2019) Sodium chloride aggravates arthritis via Th17 polarization. Yonsei Med J 60: 88-97. https://doi.org/10.3349/ymj.2019.60.1.88
|
| [59] |
Haase S, Wilck N, Kleinewietfeld M, et al. (2019) Sodium chloride triggers Th17 mediated autoimmunity. J Neuroimmunol 329: 9-13. https://doi.org/10.1016/j.jneuroim.2018.06.016
|
| [60] |
Devirgiliis C, Zinno P, Perozzi G (2013) Update on antibiotic resistance in foodborne Lactobacillus and Lactococcus species. Front Microbiol 4: 301. https://doi.org/10.3389/fmicb.2013.00301
|
Figures(1) / Tables(1)

Sayuri Yoshikawa, Kurumi Taniguchi, Haruka Sawamura, Yuka Ikeda, Ai Tsuji, Satoru Matsuda. Promising probiotics for the treatment of nephrotoxicity induced during immune-checkpoint therapy against cancers[J]. AIMS Bioengineering, 2022, 9(3): 283-292. doi: 10.3934/bioeng.2022019







 DownLoad:
DownLoad: