Citation: Svetlana N. Kovalchuk, Anna L. Arkhipova, Eugene A. Klimov. Development of real-time PCR assay for genotyping SNP rs41255693 in cattle SCD gene[J]. AIMS Agriculture and Food, 2020, 5(1): 14-19. doi: 10.3934/agrfood.2020.1.14

| [1] | Ladeira MM, Schoonmaker JP, Gionbelli MP, et al. (2016) Nutrigenomics and beef quality: A review about lipogenesis. Int J Mol Sci 17. |
| [2] |
Gebreyesus G, Buitenhuis AJ, Poulsen NA, et al. (2019) Multi-population GWAS and enrichment analyses reveal novel genomic regions and promising candidate genes underlying bovine milk fatty acid composition. BMC Genomics 20: 178. doi: 10.1186/s12864-019-5573-9

|
| [3] |
Gamarra D, Aldai N, Arakawa A, et al. (2018) Distinct correlations between lipogenic gene expression and fatty acid composition of subcutaneous fat among cattle breeds. BMC Vet Res 14: 167. doi: 10.1186/s12917-018-1481-5
|
| [4] |
Mansilla MC, Banchio CE, de Mendoza D (2008) Signalling pathways controlling fatty acid desaturation. Subcell Biochem 49: 71-99. doi: 10.1007/978-1-4020-8831-5_3
|
| [5] |
Ntambi JM (1995) The regulation of stearoyl-CoA desaturase (SCD). Prog Lipid Res 34: 139-150. doi: 10.1016/0163-7827(94)00010-J
|
| [6] | Ntambi JM (1999) Regulation of stearoyl-CoA desaturase by polyunsaturated fatty acids and cholesterol. J Lipid Res 40: 1549-1558. |
| [7] |
Weiss K, Mihaly J, Liebisch G, et al. (2011) Effect of synthetic ligands of PPAR alpha, beta/delta, gamma, RAR, RXR and LXR on the fatty acid composition of phospholipids in mice. Lipids 46: 1013-1020. doi: 10.1007/s11745-011-3593-6
|
| [8] |
Bernard L, Leroux C, Hayes H, et al. (2001) Characterization of the caprine stearoyl-CoA desaturase gene and its mRNA showing an unusually long 3'-UTR sequence arising from a single exon. Gene 281: 53-61. doi: 10.1016/S0378-1119(01)00822-8
|
| [9] |
Barton L, Kott T, Bures D, et al. (2010) The polymorphisms of stearoyl-CoA desaturase (SCD1) and sterol regulatory element binding protein-1 (SREBP-1) genes and their association with the fatty acid profile of muscle and subcutaneous fat in Fleckvieh bulls. Meat Sci 85: 15-20. doi: 10.1016/j.meatsci.2009.11.016
|
| [10] |
Chung M, Ha S, Jeong S, et al. (2000) Cloning and characterization of bovine stearoyl CoA desaturasel cDNA from adipose tissues. Biosci Biotechnol Biochem 64: 1526-1530. doi: 10.1271/bbb.64.1526
|
| [11] |
Kelsey JA, Corl BA, Collier RJ, et al. (2003) The effect of breed, parity, and stage of lactation on conjugated linoleic acid (CLA) in milk fat from dairy cows. J Dairy Sci 86: 2588-2597. doi: 10.3168/jds.S0022-0302(03)73854-5
|
| [12] |
Lawless F, Murphy JJ, Harrington D, et al. (1998) Elevation of conjugated cis-9, trans-11-octadecadienoic acid in bovine milk because of dietary supplementation. J Dairy Sci 81: 3259-3267. doi: 10.3168/jds.S0022-0302(98)75890-4
|
| [13] |
Peterson DG, Kelsey JA, Bauman DE (2002) Analysis of variation in cis-9, trans-11 conjugated linoleic acid (CLA) in milk fat of dairy cows. J Dairy Sci 85: 2164-2172. doi: 10.3168/jds.S0022-0302(02)74295-1
|
| [14] |
Milanesi E, Nicoloso L, Crepaldi P (2008) Stearoyl CoA desaturase (SCD) gene polymorphisms in Italian cattle breeds. J Anim Breed Genet 125: 63-67. doi: 10.1111/j.1439-0388.2007.00697.x
|
| [15] |
Kgwatalala PM, Ibeagha-Awemu EM, Hayes JF, et al. (2009) Stearoyl-CoA desaturase 1 3'UTR SNPs and their influence on milk fatty acid composition of Canadian Holstein cows. J Anim Breed Genet 126: 394-403. doi: 10.1111/j.1439-0388.2008.00796.x
|
| [16] | Carvajal AM, Huircan P, Dezamour JM, et al. (2016) Milk fatty acid profile is modulated by DGAT1 and SCD1 genotypes in dairy cattle on pasture and strategic supplementation. Genet Mol Res 15. |
| [17] |
Houaga I, Muigai AWT, Ng'ang'a FM, et al. (2018) Milk fatty acid variability and association with polymorphisms in SCD1 and DGAT1 genes in White Fulani and Borgou cattle breeds. Mol Biol Rep 45: 1849-1862. doi: 10.1007/s11033-018-4331-4
|
| [18] | Wathes DC, Clempson AM, Pollott GE (2012) Associations between lipid metabolism and fertility in the dairy cow. Reprod Fertil Dev 25: 48-61. |
| [19] |
Kgwatalala PM, Ibeagha-Awemu EM, Mustafa AF, et al. (2009) Stearoyl-CoA desaturase 1 genotype and stage of lactation influences milk fatty acid composition of Canadian Holstein cows. Anim Genet 40: 609-615. doi: 10.1111/j.1365-2052.2009.01887.x
|
| [20] |
Pegolo S, Cecchinato A, Mele M, et al. (2016) Effects of candidate gene polymorphisms on the detailed fatty acids profile determined by gas chromatography in bovine milk. J Dairy Sci 99: 4558-4573. doi: 10.3168/jds.2015-10420
|
| [21] | Glazko VI, Andreichenko IN, Kovalchuk SN, et al. (2017) Candidate genes for control of cattle milk production traits. Russ Agric Sci 42: 458-464. |
| [22] | Rasmussen HB (2012) Restriction fragment length polymorphism analysis of PCR-amplified fragments (PCR-RFLP) and gel electrophoresis-Valuable tool for genotyping and genetic fingerprinting. In: Magdeldin S, Gel Electrophoresis-Principles and Basics, 315-334. |
| [23] |
Babii AV, Arkhipova AL, Andreichenko IN, et al. (2018) A TaqMan PCR assay for detection of DGAT1 K232A polymorphism in cattle. AIMS Agric Food 3: 306-312. doi: 10.3934/agrfood.2018.3.306
|
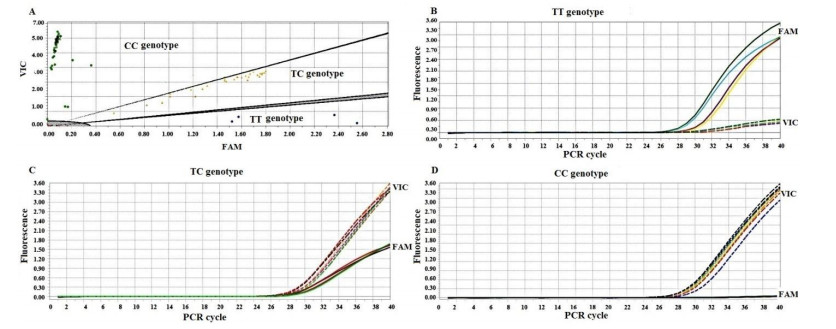
Figures(1)

Svetlana N. Kovalchuk, Anna L. Arkhipova, Eugene A. Klimov. Development of real-time PCR assay for genotyping SNP rs41255693 in cattle SCD gene[J]. AIMS Agriculture and Food, 2020, 5(1): 14-19. doi: 10.3934/agrfood.2020.1.14







 DownLoad:
DownLoad: