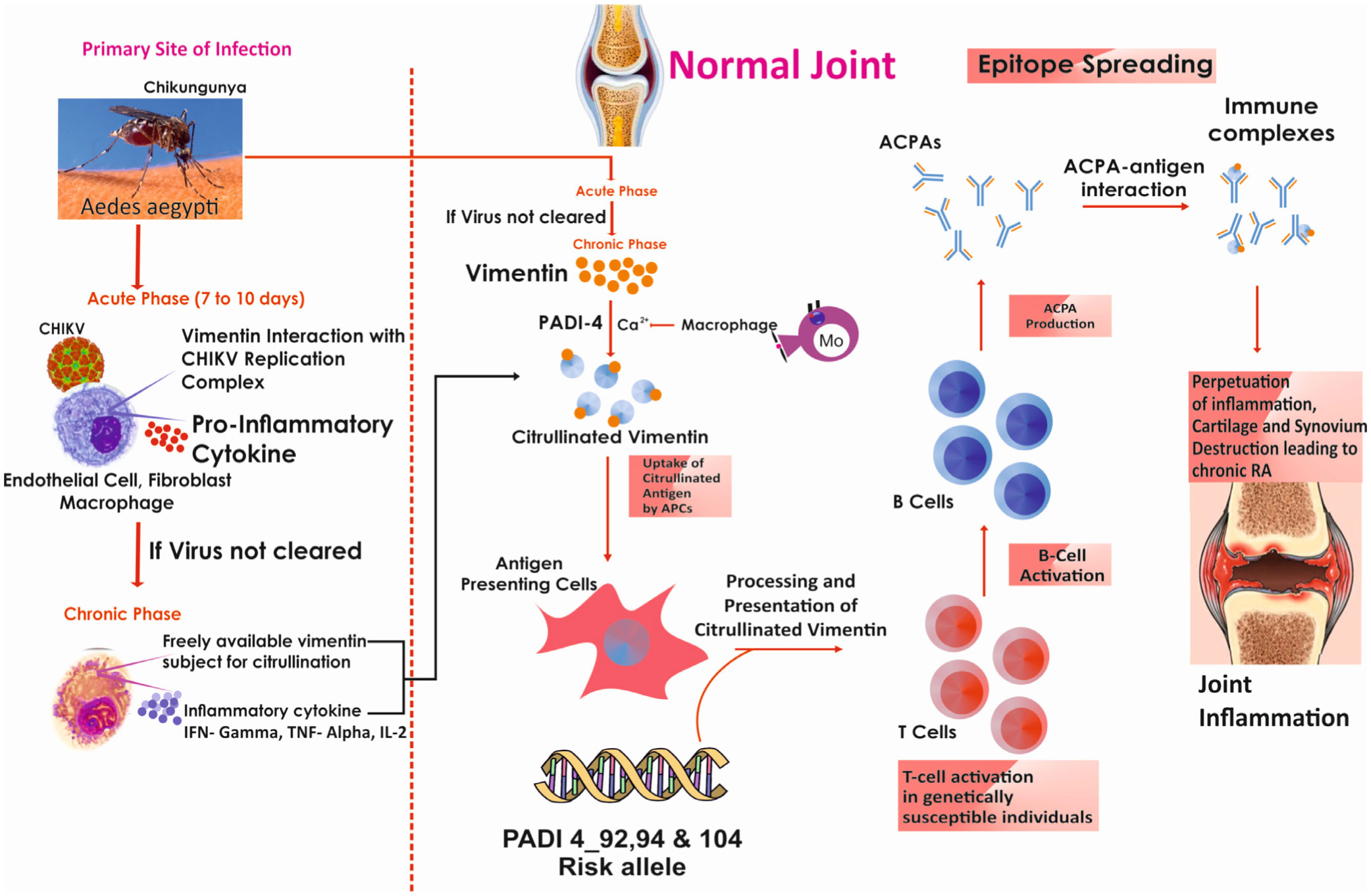
Rheumatoid arthritis (RA) is a multifactorial disease which can be triggered by gene-environment interactions. Numerous risk factors have been acknowledged in varied ethnicities, but their generalizability is vague. Hence, proposed to identify impact of etiology on citrullination and how both interact with peptidyl arginine deiminase 4 (PADI4) polymorphism in RA onset among South Indian Tamil RA cases. Studied 207 RA cases and 186 healthy controls for C-reactive protein (CRP), erythrocyte sedimentation rate (ESR), rheumatoid factor (RF), anti-cyclic citrullinated peptide (CCP), anti-Sa (citrullinated vimentin), anti-citrullinated α-enolase peptide-1 (CEP-1) and diseases activity score-28 (DAS-28). Past exposure to studied etiological risk factors obtained through questionnaire. Family history of RA (FHRA), surgery/injury and chikungunya virus (CHIKV) infection significantly contributed to RA (p < 0.05) particularly CHIKV (OR = 6.66, 95% CI 3.92–11.32, p = 0.001). Strikingly, 67.1% of surgery/injury and 80% of CHIKV exposed patients had RA onset within a year. RA cases with tooth decay had impact on RF, anti-CCP, anti-Sa freequeny and anti-CEP-1 level (p < 0.05).Since CHIKV infected cases showed significant anti-Sa (p = 0.04) level and frequency (p = 0.01), they were genotyped for polymorphism in PADI4_92 (rs874881), 104 (rs1748033) and 94 (rs2240340) by Sanger's sequencing which demonstrated that PADI4 confers risk (p < 0.05) for the onset of CHIKV induced RA. This is the initial report that CHIKV may contribute to RA development via vimentin citrullination. FHRA, surgery/injury, CHIKV and smoking posed a key RA risk.
Citation: Venkatraman Malini, Narayanasamy Shettu, Subbiah Murugesan. Impact of etiological factors on citrullination markers and susceptibility of PADI4 allele for CHIKV induced rheumatoid arthritis among South Indian Tamil RA cases[J]. AIMS Allergy and Immunology, 2022, 6(3): 153-169. doi: 10.3934/Allergy.2022012
Rheumatoid arthritis (RA) is a multifactorial disease which can be triggered by gene-environment interactions. Numerous risk factors have been acknowledged in varied ethnicities, but their generalizability is vague. Hence, proposed to identify impact of etiology on citrullination and how both interact with peptidyl arginine deiminase 4 (PADI4) polymorphism in RA onset among South Indian Tamil RA cases. Studied 207 RA cases and 186 healthy controls for C-reactive protein (CRP), erythrocyte sedimentation rate (ESR), rheumatoid factor (RF), anti-cyclic citrullinated peptide (CCP), anti-Sa (citrullinated vimentin), anti-citrullinated α-enolase peptide-1 (CEP-1) and diseases activity score-28 (DAS-28). Past exposure to studied etiological risk factors obtained through questionnaire. Family history of RA (FHRA), surgery/injury and chikungunya virus (CHIKV) infection significantly contributed to RA (p < 0.05) particularly CHIKV (OR = 6.66, 95% CI 3.92–11.32, p = 0.001). Strikingly, 67.1% of surgery/injury and 80% of CHIKV exposed patients had RA onset within a year. RA cases with tooth decay had impact on RF, anti-CCP, anti-Sa freequeny and anti-CEP-1 level (p < 0.05).Since CHIKV infected cases showed significant anti-Sa (p = 0.04) level and frequency (p = 0.01), they were genotyped for polymorphism in PADI4_92 (rs874881), 104 (rs1748033) and 94 (rs2240340) by Sanger's sequencing which demonstrated that PADI4 confers risk (p < 0.05) for the onset of CHIKV induced RA. This is the initial report that CHIKV may contribute to RA development via vimentin citrullination. FHRA, surgery/injury, CHIKV and smoking posed a key RA risk.

| [1] |
McInnes IB, Schett G (2011) The pathogenesis of rheumatoid arthritis. N Engl J Med 365: 2205-2219. https://doi.org/10.1056/NEJMra1004965 
|
| [2] |
Tobón GJ, Youinou P, Saraux A (2010) The environment, geo-epidemiology, and autoimmune disease: Rheumatoid arthritis. J Autoimmun 35: 10-14. https://doi.org/10.1016/j.jaut.2009.12.009
|
| [3] |
Cross M, Smith E, Hoy D, et al. (2014) The global burden of rheumatoid arthritis: estimates from the global burden of disease 2010 study. Ann Rheum Dis 73: 1316-1322. https://doi.org/10.1136/annrheumdis-2013-204627
|
| [4] |
van der Woude D, Alemayehu WG, Verduijn W, et al. (2010) Gene-environment interaction influences the reactivity of autoantibodies to citrullinated antigens in rheumatoid arthritis. Nat Genet 42: 814-816. https://doi.org/10.1038/ng1010-814
|
| [5] |
Deane KD, Demoruelle MK, Kelmenson LB, et al. (2017) Genetic and environmental risk factors for rheumatoid arthritis. Best Pract Res Clin Rheumatol 31: 3-18. https://doi.org/10.1016/j.berh.2017.08.003
|
| [6] |
Paul B, Pariyapurath R (2018) Risk factor assessment of rheumatoid arthritis in North Kerala. Eur J Rheumatol 5: 184-190. https://doi.org/10.5152/eurjrheum.2018.17111
|
| [7] | Brawer AE, Goel N (2016) The onset of rheumatoid arthritis following trauma. Open Access Rheumatol Res Rev 8: 77-80. https://doi.org/10.2147/OARRR.S110560 |
| [8] |
Hedström AK, Rönnelid J, Klareskog L, et al. (2019) Complex relationships of smoking, HLA-DRB1 genes, and serologic profiles in patients with early rheumatoid arthritis: update from a Swedish population-based case-control study. Arthritis Rheumatol 71: 1504-1511. https://doi.org/10.1002/art.40852
|
| [9] |
Sunar I, Garip Y, Yilmaz Ö, et al. (2015) Disease activity (rheumatoid arthritis disease activity index-5) in patients with rheumatoid arthritis and its association with quality of life, pain, fatigue, and functional and psychological status. Arch Rheumatol 30: 144-149. https://doi.org/10.5606/ArchRheumatol.2015.5122
|
| [10] |
Corbacho MI, Dapueto JJ (2010) Assessing the functional status and quality of life of patients with rheumatoid arthritis. Rev Bras Reumatol 50: 31-43. https://doi.org/10.1590/S0482-50042010000100004
|
| [11] |
Ghosh SK, Bandyopadhyay D, Biswas SK, et al. (2017) Mucocutaneous manifestations in patients with rheumatoid arthritis: A cross-sectional study from Eastern India. Indian J Dermatol 62: 411-417. https://doi.org/10.4103/ijd.IJD_260_17
|
| [12] |
Meyer PW, Ally MT, Hodkinson B, et al. (2018) Comparison of the diagnostic potential of three anti-citrullinated protein antibodies as adjuncts to rheumatoid factor and CCP in a cohort of South African rheumatoid arthritis patients. Rheumatol Int 38: 993-1001. https://doi.org/10.1007/s00296-018-4036-y
|
| [13] |
Montes A, Dieguez-Gonzalez R, Perez-Pampin E, et al. (2011) Particular association of clinical and genetic features with autoimmunity to citrullinated α-enolase in rheumatoid arthritis. Arthritis Rheum 63: 654-661. https://doi.org/10.1002/art.30186
|
| [14] | Prasanth G, Padmaraj SR, Mathew R, et al. (2013) Anti-Sa antibody, cyclic citrullinated peptide antibody and rheumatoid factor as diagnostic markers of rheumatoid arthritis. Age 212: 249-264. |
| [15] |
Mahdi H, Fisher BA, Källberg H, et al. (2009) Specific interaction between genotype, smoking and autoimmunity to citrullinated α-enolase in the etiology of rheumatoid arthritis. Nat Genet 41: 1319-1324. https://doi.org/10.1038/ng.480
|
| [16] |
Lundberg K, Kinloch A, Fisher BA, et al. (2008) Antibodies to citrullinated α-enolase peptide 1 are specific for rheumatoid arthritis and cross-react with bacterial enolase. Arthritis Rheum 58: 3009-3019. https://doi.org/10.1002/art.23936
|
| [17] |
Clowse ME, Chakravarty E, Costenbader KH, et al. (2012) Effects of infertility, pregnancy loss, and patient concerns on family size of women with rheumatoid arthritis and systemic lupus erythematosus. Arthritis Care Res 64: 668-674. https://doi.org/10.1002/acr.21593
|
| [18] |
Ramos-Remus C, Castillo-Ortiz JD, Aguilar-Lozano L, et al. (2015) Autoantibodies in prediction of the development of rheumatoid arthritis among healthy relatives of patients with the disease. Arthritis Rheumatol 67: 2837-2844. https://doi.org/10.1002/art.39297
|
| [19] |
Furman BD, Kimmerling KA, Zura RD, et al. (2015) Brief report: articular ankle fracture results in increased synovitis, synovial macrophage infiltration, and synovial fluid concentrations of inflammatory cytokines and chemokines. Arthritis Rheumatol 67: 1234-1239. https://doi.org/10.1002/art.39064
|
| [20] |
Struglics A, Larsson S, Kumahashi N, et al. (2015) Changes in cytokines and aggrecan ARGS neoepitope in synovial fluid and serum and in C-terminal crosslinking telopeptide of type II collagen and N-terminal crosslinking telopeptide of type I collagen in urine over five years after anterior cruciate ligament rupture: an exploratory analysis in the knee anterior cruciate ligament, nonsurgical versus surgical treatment trial. Arthritis Rheumatol 67: 1816-1825. https://doi.org/10.1002/art.39146
|
| [21] |
Eltzschig HK, Carmeliet P (2011) Hypoxia and inflammation. N Engl J Med 364: 656-665. https://doi.org/10.1056/NEJMra0910283
|
| [22] |
Manfredi AA, Rovere-Querini P (2010) The mitochondrion-a Trojan horse that kicks off inflammation. N Engl J Med 362: 2132-2134. https://doi.org/10.1056/NEJMcibr1003521
|
| [23] |
Swärd P, Frobell R, Englund M, et al. (2012) Cartilage and bone markers and inflammatory cytokines are increased in synovial fluid in the acute phase of knee injury (hemarthrosis)–a cross-sectional analysis. Osteoarthr Cartilage 20: 1302-1308. https://doi.org/10.1016/j.joca.2012.07.021
|
| [24] |
Snir O, Rieck M, Gebe JA, et al. (2011) Identification and functional characterization of T cells reactive to citrullinated vimentin in HLA-DRB1* 0401-positive humanized mice and rheumatoid arthritis patients. Arthritis Rheum 63: 2873-2883. https://doi.org/10.1002/art.30445
|
| [25] |
Hill JA, Southwood S, Sette A, et al. (2003) Cutting edge: the conversion of arginine to citrulline allows for a high-affinity peptide interaction with the rheumatoid arthritis-associated HLA-DRB1* 0401 MHC class II molecule. J Immunol 171: 538-541. https://doi.org/10.4049/jimmunol.171.2.538
|
| [26] |
Manimunda SP, Vijayachari P, Uppoor R, et al. (2010) Clinical progression of chikungunya fever during acute and chronic arthritic stages and the changes in joint morphology as revealed by imaging. T Roy Soc Trop Med H 104: 392-399. https://doi.org/10.1016/j.trstmh.2010.01.011
|
| [27] |
Miner JJ, Aw Yeang HX, Fox JM, et al. (2015) Brief report: chikungunya viral arthritis in the United States: a mimic of seronegative rheumatoid arthritis. Arthritis Rheumatol 67: 1214-1220. https://doi.org/10.1002/art.39027
|
| [28] |
Schilte C, Staikovsky F, Couderc T, et al. (2013) Chikungunya virus-associated long-term arthralgia: a 36-month prospective longitudinal study. PLoS Negl Trop Dis 7: e2137. https://doi.org/10.1371/journal.pntd.0002137
|
| [29] |
Chopra A, Anuradha V, Ghorpade R, et al. (2012) Acute chikungunya and persistent musculoskeletal pain following the 2006 Indian epidemic: a 2-year prospective rural community study. Epidemiol Infect 140: 842-850. https://doi.org/10.1017/S0950268811001300
|
| [30] |
Maek-a-Nantawat W, Silachamroon U (2009) Presence of autoimmune antibody in chikungunya infection. Case Rep Med 2009: 1-4. https://doi.org/10.1155/2009/840183
|
| [31] | Gauri LA, Thaned A, Fatima Q, et al. (2016) Clinical spectrum of chikungunya in Bikaner (North Western India) in 2006 and follow up of patients for five years. J Assoc Physicians India 64: 22-25. |
| [32] |
Chen W, Foo SS, Sims NA, et al. (2015) Arthritogenic alpha viruses: new insights into arthritis and bone pathology. Trends Microbiol 23: 35-43. https://doi.org/10.1016/j.tim.2014.09.005
|
| [33] |
Sugiyama D, Nishimura K, Tamaki K, et al. (2010) Impact of smoking as a risk factor for developing rheumatoid arthritis: a meta-analysis of observational studies. Ann Rheum Dis 69: 70-81. https://doi.org/10.1136/ard.2008.096487
|
| [34] |
Hedström AK, Rönnelid J, Klareskog L, et al. (2019) Complex relationships of smoking, HLA-DRB1 genes, and serologic profiles in patients with early rheumatoid arthritis: update from a Swedish population-based case–control study. Arthritis Rheumatol 71: 1504-1511. https://doi.org/10.1002/art.40852
|
| [35] | Roopa DA, Agrawal N, Johari S, et al. (2015) Prevalence of periodontitis among rheumatoid arthritis patients: An epidemiological study. Rama Univ J Dent Sci 2: 2-8. |
| [36] |
Kim JW, Park JB, Yim HW, et al. (2018) Rheumatoid arthritis is associated with early tooth loss: results from Korea National Health and Nutrition Examination Survey V to VI. Korean J Intern Med 34: 1381-1391. https://doi.org/10.3904/kjim.2018.093
|
| [37] |
Johansson K, Askling J, Alfredsson L, et al. (2018) Mediterranean diet and risk of rheumatoid arthritis: a population-based case-control study. Arthritis Res Ther 20: 1-8. https://doi.org/10.1186/s13075-018-1680-2
|
| [38] |
Hu Y, Sparks JA, Malspeis S, et al. (2017) Long-term dietary quality and risk of developing rheumatoid arthritis in women. Ann Rheum Dis 76: 1357-1364. https://doi.org/10.1136/annrheumdis-2016-210431
|
| [39] | Tsubaki M, Takeda T, Kino T, et al. (2015) Mangiferin suppresses CIA by suppressing the expression of TNF-α, IL-6, IL-1β, and RANKL through inhibiting the activation of NF-κB and ERK1/2. Am J Transl Res 7: 1371-1381. |
| [40] | Bang SY, Han TU, Choi CB, et al. (2010) Peptidyl arginine deiminase type IV (PADI4) haplotypes interact with shared epitope regardless of anti-cyclic citrullinated peptide antibody or erosive joint status in rheumatoid arthritis: a case control study. Arthritis Res Ther 12: 1-9. https://doi.org/10.1186/ar3051 |
| [41] |
Hedström AK, Stawiarz L, Klareskog L, et al. (2018) Smoking and susceptibility to rheumatoid arthritis in a Swedish population-based case-control study. Eur J Epidemiol 33: 415-423. https://doi.org/10.1007/s10654-018-0360-5
|
| [42] |
Laugisch O, Wong A, Sroka A, et al. (2016) Citrullination in the periodontium—a possible link between periodontitis and rheumatoid arthritis. Clin Oral Investig 20: 675-683. https://doi.org/10.1007/s00784-015-1556-7
|
| [43] |
Konig MF, Abusleme L, Reinholdt J, et al. (2016) Aggregatibacter actinomycetemcomitans-induced hypercitrullination links periodontal infection to autoimmunity in rheumatoid arthritis. Sci Transl Med 8: 369ra176. https://doi.org/10.1126/scitranslmed.aaj1921
|
| [44] |
Wegner N, Wait R, Sroka A, et al. (2010) Peptidyl arginine deiminase from Porphyromonasgingivalis citrullinates human fibrinogen and α-enolase: Implications for autoimmunity in rheumatoid arthritis. Arthritis Rheum 62: 2662-2672. https://doi.org/10.1002/art.27552
|
| [45] |
Issac THK, Tan EL, Chu JJH (2014) Proteomic profiling of chikungunya virus-infected human muscle cells: reveal the role of cytoskeleton network in CHIKV replication. J Proteomics 108: 445-464. https://doi.org/10.1016/j.jprot.2014.06.003
|
| [46] |
Fox JM, Diamond MS (2016) Immune-mediated protection and pathogenesis of chikungunya virus. J Immunol 197: 4210-4218. https://doi.org/10.4049/jimmunol.1601426
|
| [47] |
Phuklia W, Kasisith J, Modhiran N, et al. (2013) Osteoclastogenesis induced by CHIKV-infected fibroblast-like synoviocytes: a possible interplay between synoviocytes and monocytes/macrophages in CHIKV-induced arthralgia/arthritis. Virus Res 177: 179-188. https://doi.org/10.1016/j.virusres.2013.08.011
|
| [48] |
Wauquier N, Becquart P, Nkoghe D, et al. (2010) The acute phase of Chikungunya virus infection in humans is associated with strong innate immunity and T CD8 cell activation. J Infect Dis 204: 115-123. https://doi.org/10.1093/infdis/jiq006
|
| [49] |
Hoarau JJ, Bandjee MCJ, Trotot PK, et al. (2010) Persistent chronic inflammation and infection by chikungunya arthritogenic alphavirus in spite of a robust host immune response. J Immunol 184: 5914-5927. https://doi.org/10.4049/jimmunol.0900255
|
| [50] |
Vossenaar ER, Després N, Lapointe E, et al. (2004) Rheumatoid arthritis specific anti-Sa antibodies target citrullinated vimentin. Arthritis Res Ther 6: R142-R150. https://doi.org/10.1186/ar1057
|
Figures(1) / Tables(6)

Venkatraman Malini, Narayanasamy Shettu, Subbiah Murugesan. Impact of etiological factors on citrullination markers and susceptibility of PADI4 allele for CHIKV induced rheumatoid arthritis among South Indian Tamil RA cases[J]. AIMS Allergy and Immunology, 2022, 6(3): 153-169. doi: 10.3934/Allergy.2022012







 DownLoad:
DownLoad: