Citation: Albert Uchenna Ude, Che Husna Azhari. Crashworthiness response of natural silk-fibre glass hybrid reinforced epoxy cylindrical composite tubes under quasi-static load[J]. AIMS Materials Science, 2019, 6(5): 852-863. doi: 10.3934/matersci.2019.5.852

| [1] |
Ude A, Azhari C (2013) Experimental investigation on the response of woven natural silk fiber/epoxy sndwich composite panels under low velocity impact. Fiber Polym 14: 127-132. doi: 10.1007/s12221-013-0127-2

|
| [2] |
Eshkoor R, Ude A, Oshkovr S, et al. (2014) Failure mechanism of woven natural silk/epoxy rectangular composite tubes under axial quasi-static crushing test using trigger mechanism. Int J Impact Eng 64: 53-61. doi: 10.1016/j.ijimpeng.2013.09.004
|
| [3] |
Eshkoor R, Ude A, Sulong A, et al. (2015) Energy absorption and load carrying capability of woven natural silk epoxy-triggered composite tubes. Compos Part B-Eng 77: 10-18. doi: 10.1016/j.compositesb.2015.03.017
|
| [4] |
Ude A, Ariffin A, Azhari C (2013) Impact damage characteristics in reinforced woven natural silk/epoxy composite face-sheet and sandwich foam, coremat and honeycomb materials. Int J Impact Eng 58: 31-38. doi: 10.1016/j.ijimpeng.2013.03.003
|
| [5] |
Eshkoor R, Oshkovr S, Sulong A, et al. (2013) Comparative research on the crashworthiness characteristics of woven natural silk/epoxy composite tubes. Mater Design 47: 248-257. doi: 10.1016/j.matdes.2012.11.030
|
| [6] |
Eshkoor R, Oshkovr S, Sulong A, et al. (2013) Effect of trigger configuration on the crashworthiness characteristics of natural silk epoxy composite tubes. Compos Part B-Eng 55: 5-10. doi: 10.1016/j.compositesb.2013.05.022
|
| [7] |
Zhou J, Guan Z, Cantwell W (2018) The energy-absorbing behaviour of composite tubereinforced foams. Compos Part B-Eng 139: 227-237. doi: 10.1016/j.compositesb.2017.11.066
|
| [8] |
Alkbir M, Sapuan S, Nuraini A, et al. (2016) Fibre properties and crashworthiness parameters of natural fibre-reinforced composite structure: A literature review. Compos Struct 148: 59-73. doi: 10.1016/j.compstruct.2016.01.098
|
| [9] |
Abdewi EF, Sulaiman S, Hamouda AMS, et al. (2008) Quasi-static axial and lateral crushing of radial corrugated composite tubes. Thin Wall Struct 46: 320-332. doi: 10.1016/j.tws.2007.07.018
|
| [10] | Abosbaia A, Mahdi E, Hamouda A, et al. (2008) Energy absorption capability of laterally loaded segmented composite tubes. Compos Struct 70: 356-373. |
| [11] |
Duarte I, Vesenjak M, Krstulović-Opara L, et al. (2015) Static and dynamic axial crush performance of in-situ foam-filled tubes. Compos Struct 124: 128-139. doi: 10.1016/j.compstruct.2015.01.014
|
| [12] |
Kim H, Shin D, Lee J, et al. (2014) Crashworthiness of aluminum/CFRP square hollow section beam under axial impact loading for crash box application. Compos Struct 112: 1-10. doi: 10.1016/j.compstruct.2014.01.042
|
| [13] |
Liu Q, Xing H, Ju Y, et al. (2014) Quasi-static axial crushing and transverse bending of double hat shaped CFRP tubes. Compos Struct 117: 1-11. doi: 10.1016/j.compstruct.2014.06.024
|
| [14] |
Mahdi E, Hamouda A, Sen A (2004) Quasi-static crushing behaviour of hybrid and nonhybrid natural fibre composite solid cones. Compos Struct 66: 647-663. doi: 10.1016/j.compstruct.2004.06.001
|
| [15] |
Mamalis A, Manolakos D, Ioannidis M, et al. (2009) On the crashworthiness of composite rectangular thin-walled tubes internally reinforced with aluminium or polymeric foams: Experimental and numerical simulation. Compos Struct 89: 416-423. doi: 10.1016/j.compstruct.2008.09.008
|
| [16] | Mamalis A, Manolakos D, Ioannidis M, et al. (2005) Crashworthy characteristics of axially statically compressed thin-walled square CFRP composite tubes: experimental. Compos Struct 63: 347-360. |
| [17] |
Oshkovr S, Eshkoor R, Taher S, et al. (2012) Crashworthiness characteristics investigation of silk/epoxy composite square tubes. Compos Struct 94: 2337-2342. doi: 10.1016/j.compstruct.2012.03.031
|
| [18] |
Striewe J, Reuter C, Sauerland K, et al. (2018) Manufacturing and crashworthiness of fabric-reinforced thermoplastic composites. Thin Wall Struct 123: 501-508. doi: 10.1016/j.tws.2017.11.011
|
| [19] |
Kathiresan M, Manisekar K (2017) Low velocity axial collapse behavior of E-glass fiber/epoxy composite conical frusta. Compos Struct 166: 1-11. doi: 10.1016/j.compstruct.2017.01.041
|
| [20] |
Zhang Z, Sun W, Zhao Y, et al. (2018) Crashworthiness of different composite tubes by experiments and simulations. Compos Part B-Eng 143: 86-95. doi: 10.1016/j.compositesb.2018.01.021
|
| [21] |
Mamalis A, Manolakos D, Ioannidis M, et al. (2005) On the response of thin-walled CFRP composite tubular components subjected to static and dynamic axial compressive loading: experimental. Compos Struct 69: 407-420. doi: 10.1016/j.compstruct.2004.07.021
|
| [22] |
Mamalis A, Robinson M, Manolakos D, et al. (1997) Crashworthy capability of composite material structures. Compos Struct 37: 109-134. doi: 10.1016/S0263-8223(97)80005-0
|
| [23] |
Oshkovr S, Taher S, Oshkour A, et al. (2013) Finite element modelling of axially crushed silk/epoxy composite square tubes. Compos Struct 95: 411-418. doi: 10.1016/j.compstruct.2012.07.032
|
| [24] |
Ude A, Eshkoor R, Azhari C (2017) Crashworthy characteristics of axial quasi-statically compressed Bombyx mori composite cylindrical tubes: experimental. Fiber Polym 18: 1594-1601. doi: 10.1007/s12221-017-1235-1
|
| [25] | Supian A, Sapuan S, Zuhri M, et al. (2018) Hybrid reinforced thermoset polymer composite in energy absorption tube application: A review. Def Technol 281: 112-118. |
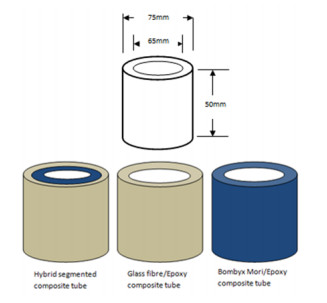
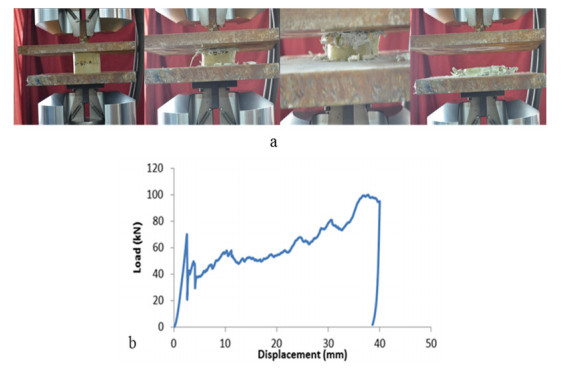
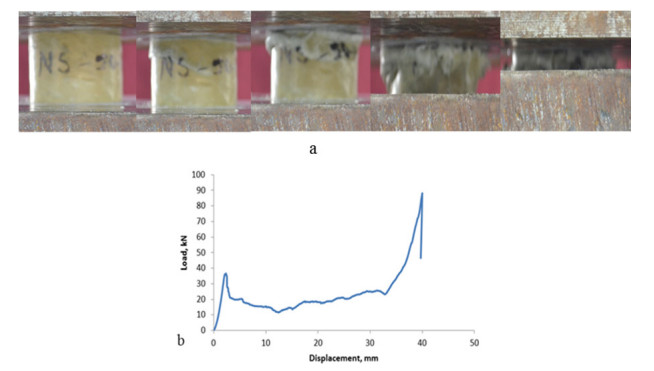
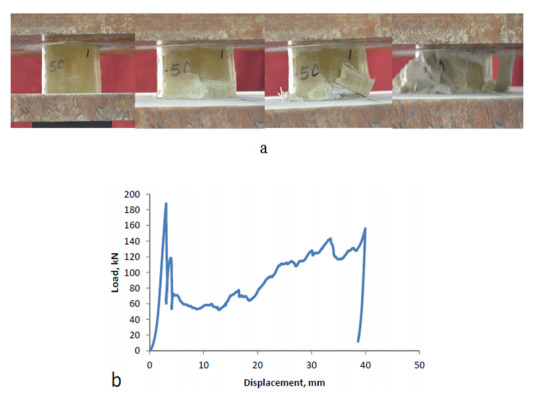

Figures(9)

Albert Uchenna Ude, Che Husna Azhari. Crashworthiness response of natural silk-fibre glass hybrid reinforced epoxy cylindrical composite tubes under quasi-static load[J]. AIMS Materials Science, 2019, 6(5): 852-863. doi: 10.3934/matersci.2019.5.852







 DownLoad:
DownLoad: