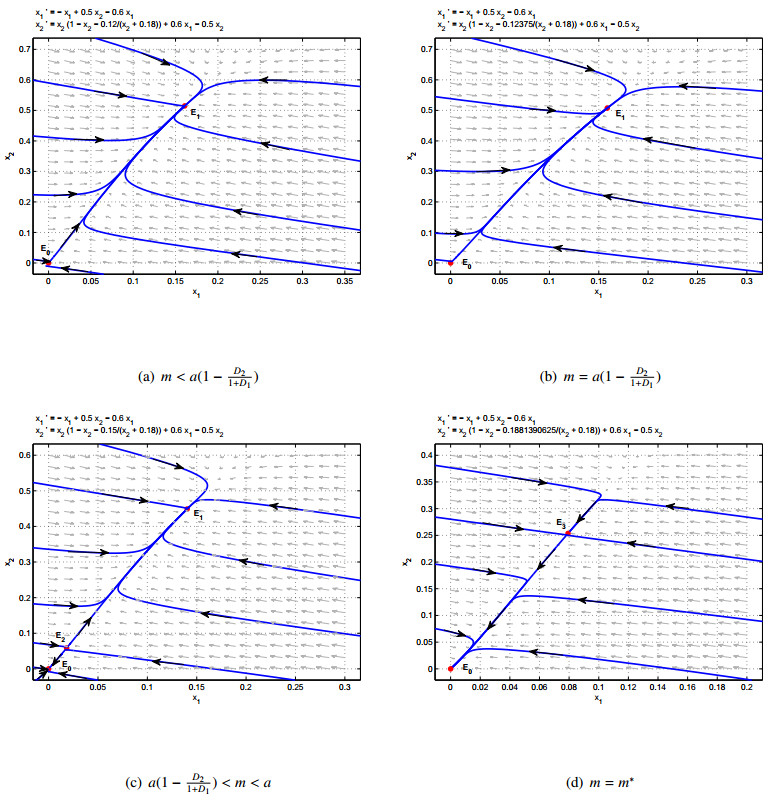
A two-patch model with additive Allee effect is proposed and studied in this paper. Our objective is to investigate how dispersal and additive Allee effect have an impact on the above model's dynamical behaviours. We discuss the local and global asymptotic stability of equilibria and the existence of the saddle-node bifurcation. Complete qualitative analysis on the model demonstrates that dispersal and Allee effect may lead to persistence or extinction in both patches. Also, combining mathematical analysis with numerical simulation, we verify that the total population abundance will increase when the Allee effect constant $ a $ increases or $ m $ decreases. And the total population density increases when the dispersal rate $ D_{1} $ increases or the dispersal rate $ D_{2} $ decreases.
Citation: Lijuan Chen, Tingting Liu, Fengde Chen. Stability and bifurcation in a two-patch model with additive Allee effect[J]. AIMS Mathematics, 2022, 7(1): 536-551. doi: 10.3934/math.2022034
A two-patch model with additive Allee effect is proposed and studied in this paper. Our objective is to investigate how dispersal and additive Allee effect have an impact on the above model's dynamical behaviours. We discuss the local and global asymptotic stability of equilibria and the existence of the saddle-node bifurcation. Complete qualitative analysis on the model demonstrates that dispersal and Allee effect may lead to persistence or extinction in both patches. Also, combining mathematical analysis with numerical simulation, we verify that the total population abundance will increase when the Allee effect constant $ a $ increases or $ m $ decreases. And the total population density increases when the dispersal rate $ D_{1} $ increases or the dispersal rate $ D_{2} $ decreases.

| [1] |
H. Andren, Effects of habitat fragmentation on birds and mammals in landscapes with different proportions of suitable habitat: A review, Oikos, 71 (1994), 355–366. doi: 10.2307/3545823. doi: 10.2307/3545823

|
| [2] |
J. G. Skellam, Random dispersal in theoretical populations, Biometrika, 38 (1951), 196–218. doi: 10.2307/2332328. doi: 10.2307/2332328
|
| [3] |
H. I. Freedman, P. Waltman, Mathematical models of population interactions with dispersal. I: Stability of two habitats with and without a predator, SIAM J. App. Math., 32 (1977), 631–648. doi: 10.1137/0132052. doi: 10.1137/0132052
|
| [4] |
D. L. DeAngelis, C. C. Travis, W. M. Post, Persistence and stability of seed-dispersed species in a patchy environment, Theor. Popul. Biol., 16 (1979), 107–125. doi: 10.1016/0040-5809(79)90008-X. doi: 10.1016/0040-5809(79)90008-X
|
| [5] |
R. D. Holt, Population dynamics in two-patch environments: Some anomalous consequences of an optimal habitat distribution, Theor. Popul. Biol., 28 (1985), 181–208. doi: 10.1016/0040-5809(85)90027-9. doi: 10.1016/0040-5809(85)90027-9
|
| [6] |
D. L. DeAngelis, B. Zhang, Effects of dispersal in a non-uniform environment on population dynamics and competition: A patch model approach, Discrete Cont. Dyn-A., 19 (2014), 3087–3104. doi: 10.3934/dcdsb.2014.19.3087. doi: 10.3934/dcdsb.2014.19.3087
|
| [7] |
A. Roger, L. Claude, S. Tewfik, Is dispersal always beneficial to carrying capacity? New insights from the multi-patch logistic equation, Theor. Popul. Biol., 106 (2015), 45–59. doi: 10.1016/j.tpb.2015.10.001. doi: 10.1016/j.tpb.2015.10.001
|
| [8] |
R. Arditi, C. Lobry, T. Sari, Asymmetric dispersal in the multi-patch logistic equation, Theor. Popul. Biol., 120 (2018), 11–15. doi: 10.1016/j.tpb.2017.12.006. doi: 10.1016/j.tpb.2017.12.006
|
| [9] |
H. Wu, Y. S. Wang, Y. F. Li, D. L. DeAngelis, Dispersal asymmetry in a two-patch system with source-sink populations, Theor. Popul. Biol., 131 (2020), 54–65. doi: 10.1016/j.tpb.2019.11.004. doi: 10.1016/j.tpb.2019.11.004
|
| [10] |
R. Huang, Y. S. Wang, H. Wu, Population abundance in predator-prey systems with predator's dispersal between two patches, Theor. Popul. Biol., 135 (2020), 1–8. doi: 10.1016/j.tpb.2020.06.002. doi: 10.1016/j.tpb.2020.06.002
|
| [11] |
M. Fang, Y. Wang, M. Chen, L. DeAngelis, Asymptotic population abundance of a two-patch system with asymmetric diffusion, Discrete Cont. Dyn-A., 40 (2020), 3411–3425. doi: 10.3934/dcds.2020031. doi: 10.3934/dcds.2020031
|
| [12] |
P. A. Stephens, W. J. Sutherland, R. P. Freckleton, What is the Allee effect, Oikos, 87 (1999), 185–190. doi: 10.2307/3547011. doi: 10.2307/3547011
|
| [13] |
P. M. Hao, X. C. Wang, J. J. Wei, Hopf bifurcation analysis of a diffusive single species model with stage structure and strong Allee effect, Math. Comput. Simulat., 153 (2018), 1–14. doi: 10.1016/j.matcom.2018.05.004. doi: 10.1016/j.matcom.2018.05.004
|
| [14] |
L. M. Zhang, C. F. Zhang, Z. R. He, Codimension-one and codimension-two bifurcations of a discrete predator-prey system with strong Allee effect, Math. Comput. Simulat., 162 (2019), 155–178. doi: 10.1016/j.matcom.2019.01.006. doi: 10.1016/j.matcom.2019.01.006
|
| [15] |
C. Arancibia-Ibarra, J. Flores, Dynamics of a Leslie-Gower predator-prey model with Holling type II functional response, Allee effect and a generalist predator, Math. Comput. Simulat., 188 (2021), 1–12. doi: 10.1016/j.matcom.2021.03.035. doi: 10.1016/j.matcom.2021.03.035
|
| [16] |
Y. Kang, N. Lanchier, Expansion or extinction: Deterministic and stochastic two-patch models with Allee effects, J. Math. Biol., 62 (2011), 925–973. doi: 10.1007/s00285-010-0359-3. doi: 10.1007/s00285-010-0359-3
|
| [17] |
D. Pal, G. P. Samanta, Effects of dispersal speed and strong Allee effect on stability of a two-patch predator-prey model, Int. J. Dyn. Control, 6 (2018), 1484–1495. doi: 10.1007/s40435-018-0407-1. doi: 10.1007/s40435-018-0407-1
|
| [18] |
S. Saha, G. P. Samanta, Influence of dispersal and strong Allee effect on a two-patch predator-prey model, Int. J. Dyn. Control, 7 (2019), 1321–1349. doi: 10.1007/s40435-018-0490-3. doi: 10.1007/s40435-018-0490-3
|
| [19] |
B. Dennis, Allee effects: Population growth, critical density and the chance of extinction, Nat. Resour. Model, 3 (1989), 481–538. doi: 10.1111/j.1939-7445.1989.tb00119.x. doi: 10.1111/j.1939-7445.1989.tb00119.x
|
| [20] |
Y. Y. Lv, L. J. Chen, F. D. Chen, Stability and bifurcation in a single species logistic model with additive Allee effect and feedback control, Adv. Differ. Equ., 8 (2020), 2686–2697. doi: 10.1186/s13662-020-02586-0. doi: 10.1186/s13662-020-02586-0
|
| [21] |
Y. Y. Lv, L. J. Chen, F. D. Chen, Z. Li, Stability and bifurcation in an SI epidemic model with additive Allee effect and time delay, Int. J. Bifurcat. Chaos, 31 (2021), 2150060. doi: 10.1142/S0218127421500607. doi: 10.1142/S0218127421500607
|
| [22] |
J. Y. Xu, T. H. Zhang, M. A. Han, A regime switching model for species subject to environmental noises and additive Allee effect, Physica A, 527 (2019), 121300. doi: 10.1016/j.physa.2019.121300. doi: 10.1016/j.physa.2019.121300
|
| [23] |
Y. L. Cai, C. D. Zhao, W. M. Wang, J. F. Wang, Dynamics of a Leslie-Gower predator-prey model with additive Allee effect, App. Math. Model., 39 (2015), 2092–2106. doi: 10.1016/j.apm.2014.09.038. doi: 10.1016/j.apm.2014.09.038
|
| [24] |
W. D. Wang, Population dispersal and Allee effect, Ric. Mat., 65 (2016), 535–548. doi: 10.1007/s11587-016-0273-0. doi: 10.1007/s11587-016-0273-0
|
| [25] | Z. F. Zhang, T. R. Ding, W. Z. Huang, Z. X. Dong, Qualitative Theory of Differential Equation, Bei Jing: Science Press, 1992. doi: 10.1007/978-1-4757-4969-4_4. |
| [26] | L. Perko, Differential Equations and Dynamical Systems, 3 Eds., New York: Springer-Verlag, 2001. doi: 10.1007/978-1-4613-0003-8. |
| [27] |
M. Gazor, M. Kazemi, Singularity: A maple library for local zero bifurcation control of scalar smooth maps, ASME J. Comput. Nonlinear Dyn., 15 (2020), 011001. doi: 10.1115/1.4045286. doi: 10.1115/1.4045286
|
| [28] |
M. Gazor, M. Kazemi, Normal Form Analysis of Z$_2$-Equivariant Singularities, Int. J. Bifurcat. Chaos, 29 (2019), 1950015. doi: 10.1142/S0218127419500159. doi: 10.1142/S0218127419500159
|
| [29] |
B. W. Qin, K. W. Chung, A. Algaba, A. J. Rodríguez-Luis, High-order approximation of heteroclinic bifurcations in truncated 2D-normal forms for the generic cases of Hopf-zero and nonresonant double Hopf singularities, SIAM J. Appl. Dyn. Syst., 20 (2021), 403–437. doi: 10.1137/20M1325101. doi: 10.1137/20M1325101
|
| [30] |
M. Gazor, N. Sadri, Bifurcation controller designs for the generalized cusp plants of Bogdanov–Takens singularity with an application to ship control, SIAM J. Control Optim., 57 (2019), 2122–2151. doi: 10.1137/18M1210769. doi: 10.1137/18M1210769
|
| [31] |
M. Gazor, N. Sadri, Bifurcation control and universal unfolding for Hopf-zero singularities with Leading solenoidal terms, SIAM J. Appl. Dyn. Syst., 15 (2016), 870–903. doi: 10.1137/141000403. doi: 10.1137/141000403
|
| [32] |
A. J. Krener, W. Kang, D. E. Chang, Control bifurcations, IEEE T. Automat. Contr., 49 (2004), 1231–1246. doi: 10.1109/TAC.2004.832199. doi: 10.1109/TAC.2004.832199
|
| [33] |
B. Hamzi, W. Kang, J. P. Barbot, Analysis and control of Hopf bifurcations, SIAM J. Control Optim., 42 (2004), 2200–2220. doi: 10.1137/S0363012900372714. doi: 10.1137/S0363012900372714
|



Figures(4)

Lijuan Chen, Tingting Liu, Fengde Chen. Stability and bifurcation in a two-patch model with additive Allee effect[J]. AIMS Mathematics, 2022, 7(1): 536-551. doi: 10.3934/math.2022034







 DownLoad:
DownLoad: