This paper investigates the complex dynamical behavior of a discrete prey-predator system with a fear factor, a strong Allee effect, and prey refuge. The existence and stability of fixed points in the system are discussed. By applying the central manifold theorem and bifurcation theory, we have established the occurrence of various types of bifurcations, including flip bifurcation and Neimark-Sacker bifurcation. Furthermore, to address the observed chaotic behavior in the system, three controllers were designed by employing state feedback control, OGY feedback control, and hybrid control methods. These controllers serve to control chaos in the proposed system and identify specific conditions under which chaos or bifurcations can be stabilized. Finally, the theoretical analyses have been validated through numerical simulations.
Citation: Xiaoming Su, Jiahui Wang, Adiya Bao. Stability analysis and chaos control in a discrete predator-prey system with Allee effect, fear effect, and refuge[J]. AIMS Mathematics, 2024, 9(5): 13462-13491. doi: 10.3934/math.2024656
This paper investigates the complex dynamical behavior of a discrete prey-predator system with a fear factor, a strong Allee effect, and prey refuge. The existence and stability of fixed points in the system are discussed. By applying the central manifold theorem and bifurcation theory, we have established the occurrence of various types of bifurcations, including flip bifurcation and Neimark-Sacker bifurcation. Furthermore, to address the observed chaotic behavior in the system, three controllers were designed by employing state feedback control, OGY feedback control, and hybrid control methods. These controllers serve to control chaos in the proposed system and identify specific conditions under which chaos or bifurcations can be stabilized. Finally, the theoretical analyses have been validated through numerical simulations.

| [1] |
Y. Nutku, Hamiltonian structure of the Lotka-Volterra equations, Phys. Lett. A, 145 (1990), 27–28. https://doi.org/10.1016/0375-9601(90)90270-X doi: 10.1016/0375-9601(90)90270-X

|
| [2] |
M. Haque, A predator-prey model with disease in the predator species only, Nonlinear Anal.-Real, 11 (2010), 2224–2236. https://doi.org/10.1016/j.nonrwa.2009.06.012 doi: 10.1016/j.nonrwa.2009.06.012
|
| [3] |
M. Haque, A detailed study of the Beddington-DeAngelis predator-prey model, Math. Biosci., 234 (2011), 1–16. https://doi.org/10.1016/j.mbs.2011.07.003 doi: 10.1016/j.mbs.2011.07.003
|
| [4] |
P. Majumdar, B. Mondal, S. Debnathm, U. Ghosh, Controlling of periodicity and chaos in a three dimensional prey predator model introducing the memory effect, Chaos Soliton. Fract., 164 (2022), 112585. https://doi.org/10.1016/j.chaos.2022.112585 doi: 10.1016/j.chaos.2022.112585
|
| [5] |
Q. L. Chen, Z. D. Teng, Z. Y. Hu, Bifurcation and control for a discrete-time prey-predator model with Holling-Ⅳ functional response, Int. J. Ap. Mat. Com.-Pol., 23 (2013), 247–261. https://doi.org/10.2478/amcs-2013-0019 doi: 10.2478/amcs-2013-0019
|
| [6] |
J. L. Chen, Z. L. Zhu, X. Q. He, F. D. Chen, Bifurcation and chaos in a discrete predator-prey system of Leslie type with Michaelis-Menten prey harvesting, Open Math., 20 (2022), 608–628. https://doi.org/10.1515/math-2022-0054 doi: 10.1515/math-2022-0054
|
| [7] |
J. Roy, S. Dey, M. Banerjee, Maturation delay induced stability enhancement and shift of bifurcation thresholds in a predator-prey model with generalist predator, Math. Comput. Simulat., 211 (2023), 368–393. https://doi.org/10.1016/j.matcom.2023.04.019 doi: 10.1016/j.matcom.2023.04.019
|
| [8] | W. C. Allee, Animal aggregations: A study in general sociology, Chicago: University of Chicago Press, 1931. https://doi.org/10.5962/bhl.title.7313 |
| [9] |
P. A. Stephens, W. J. Sutherland, Consequences of the Allee effect for behaviour, ecology and conservation, Trends Ecol. Evol., 14 (1999), 401–405. https://doi.org/10.1016/S0169-5347(99)01684-5 doi: 10.1016/S0169-5347(99)01684-5
|
| [10] |
E. Angulo, G. M. Luque, S. D. Gregory, J. W. Wenzel, C. B. Gomes, L. Berec, et al., Allee effects in social species, J. Anim. Ecol., 87 (2018), 47–58. https://doi.org/10.1111/1365-2656.12759 doi: 10.1111/1365-2656.12759
|
| [11] |
W. Z. Lidicker Jr, The Allee effect: Its history and future importance, Open Ecol. J., 3 (2010), 71–82. https://doi.org/10.2174/1874213001003010071 doi: 10.2174/1874213001003010071
|
| [12] |
A. Arsie, C. Kottegoda, C. Shan, A predator-prey system with generalized Holling type Ⅳ functional response and Allee effects in prey, J. Differ. Equations, 309 (2022), 704–740. https://doi.org/10.1016/j.jde.2021.11.041 doi: 10.1016/j.jde.2021.11.041
|
| [13] |
L. M. Zhang, T. Wang, Qualitative properties, bifurcations and chaos of a discrete predator-prey system with weak Allee effect on the predator, Chaos Soliton. Fract., 175 (2023), 113995. https://doi.org/10.1016/j.chaos.2023.113995 doi: 10.1016/j.chaos.2023.113995
|
| [14] |
Y. Chang, W. Feng, M. Freeze, X. Lu, C. Smith, Elimination, permanence, and exclusion in a competition model under Allee effects, AIMS Math., 8 (2023), 7787–7805. https://doi.org/10.3934/math.2023391 doi: 10.3934/math.2023391
|
| [15] |
S. Creel, D. Christianson, S. Liley, J. A. Winnie Jr, Predation risk affects reproductive physiology and demography of elk, Science, 315 (2007), 960–960. https://doi.org/10.1126/science.1135918 doi: 10.1126/science.1135918
|
| [16] |
E. L. Preisser, D. I. Bolnick, The many faces of fear: Comparing the pathways and impacts of nonconsumptive predator effects on prey populations, PloS One, 3 (2008). https://doi.org/10.1371/journal.pone.0002465 doi: 10.1371/journal.pone.0002465
|
| [17] |
K. B. Altendorf, J. W. Laundré, C. A. L. González, J. S. Brown, Assessing effects of predation risk on foraging behavior of mule deer, J. Mammal., 82 (2001), 430–439. https://doi.org/10.1644/1545-1542(2001)082<0430:AEOPRO>2.0.CO;2 doi: 10.1644/1545-1542(2001)082<0430:AEOPRO>2.0.CO;2
|
| [18] |
S. Creel, D. Christianson, Relationships between direct predation and risk effects, Trends Ecol. Evol., 23 (2008), 194–201. https://doi.org/10.1016/j.tree.2007.12.004 doi: 10.1016/j.tree.2007.12.004
|
| [19] |
S. L. Lima, Nonlethal effects in the ecology of predator-prey interactions, Bioscience, 48 (1998), 25–34. https://doi.org/10.2307/1313225 doi: 10.2307/1313225
|
| [20] |
S. L. Lima, Predators and the breeding bird: Behavioral and reproductive flexibility under the risk of predation, Biol. Rev., 84 (2009), 485–513. https://doi.org/10.1111/j.1469-185X.2009.00085.x doi: 10.1111/j.1469-185X.2009.00085.x
|
| [21] |
Z. W. Liang, X. Y. Meng, Stability and Hopf bifurcation of a multiple delayed predator-prey system with fear effect, prey refuge and Crowley-Martin function, Chaos Soliton. Fract., 175 (2023), 113955. https://doi.org/10.1016/j.chaos.2023.113955 doi: 10.1016/j.chaos.2023.113955
|
| [22] |
S. Debnath, P. Majumdar, S. Sarkar, U. Ghosh, Memory effect on prey-predator dynamics: Exploring the role of fear effect, additional food and anti-predator behaviour of prey, J. Comput. Sci., 55 (2023), 101929. https://doi.org/10.1016/j.jocs.2022.101929 doi: 10.1016/j.jocs.2022.101929
|
| [23] |
B. F. Xie, N. Zhang, Influence of fear effect on a Holling type Ⅲ prey-predator system with the prey refuge, AIMS Math., 7 (2022), 1811–1830. https://doi.org/10.3934/math.2022104 doi: 10.3934/math.2022104
|
| [24] |
J. N. McNair, The effects of refuges on predator-prey interactions: A reconsideration, Theor. Popul. Biol., 29 (1986), 38–63. https://doi.org/10.1016/0040-5809(86)90004-3 doi: 10.1016/0040-5809(86)90004-3
|
| [25] |
A. Sih, Prey refuges and predator-prey stability, Theor. Popul. Biol., 31 (1987), 1–12. https://doi.org/10.1016/0040-5809(87)90019-0 doi: 10.1016/0040-5809(87)90019-0
|
| [26] |
V. Krivan, Effects of optimal antipredator behavior of prey on predator-prey dynamics: The role of refuges, Theor. Popul. Biol., 53 (1998), 131–142. https://doi.org/10.1006/tpbi.1998.1351 doi: 10.1006/tpbi.1998.1351
|
| [27] |
A. C. Crombie, Further experiments on insect competition, P. Roy. Soc. London S., 133 (1946), 76–109. https://doi.org/10.1098/rspb.1946.0004 doi: 10.1098/rspb.1946.0004
|
| [28] |
J. Ghosh, B. Sahoo, S. Poria, Prey-predator dynamics with prey refuge providing additional food to predator, Chaos Soliton. Fract., 96 (2017), 110–119. https://doi.org/10.1016/j.chaos.2017.01.010 doi: 10.1016/j.chaos.2017.01.010
|
| [29] |
A. Das, G. P. Samanta, A prey-predator model with refuge for prey and additional food for predator in a fluctuating environment, Phys. A, 538 (2020), 1228449. https://doi.org/10.1016/j.physa.2019.122844 doi: 10.1016/j.physa.2019.122844
|
| [30] |
A. A. Thirthar, S. J. Majeed, M. A. Alqudah, P. Panja, T. Abdeljawad, Fear effect in a predator-prey model with additional food, prey refuge and harvesting on super predator, Chaos Soliton. Fract., 159 (2022), 112091. https://doi.org/10.1016/j.chaos.2022.112091 doi: 10.1016/j.chaos.2022.112091
|
| [31] |
K. P. Hadeler, H. Gerstmann, The discrete Rosenzweig model, Math. Biosci., 98 (1990), 49–72. https://doi.org/10.1016/0025-5564(90)90011-M doi: 10.1016/0025-5564(90)90011-M
|
| [32] |
M. B. Ajaz, U. Saeed, Q. Din, I. Ali, I. M. I. Siddiqui, Bifurcation analysis and chaos control in discrete-time modified Leslie-Gower prey harvesting model, Adv. Differential Equ., 2020 (2020), 1–24. https://doi.org/10.1186/s13662-020-2498-1 doi: 10.1186/s13662-020-2498-1
|
| [33] |
P. A. Naik, Z. Eskandari, M. Yavuz, J. Zu, Complex dynamics of a discrete-time Bazykin-Berezovskaya prey-predator model with a strong Allee effect, J. Comput. Appl. Math., 413 (2022), 114401. https://doi.org/10.1016/j.cam.2022.114401 doi: 10.1016/j.cam.2022.114401
|
| [34] |
T. W. Zhang, Y. T. Liu, H. Z. Qu, Global mean-square exponential stability and random periodicity of discrete-time stochastic inertial neural networks with discrete spatial diffusions and Dirichlet boundary condition, Comput. Math. Appl., 141 (2023), 116–128. https://doi.org/10.1016/j.camwa.2023.04.011 doi: 10.1016/j.camwa.2023.04.011
|
| [35] |
T. W. Zhang, Z. H. Li, Switching clusters' synchronization for discrete space-time complex dynamical networks via boundary feedback controls, Pattern Recogn., 143 (2023), 109763. https://doi.org/10.1016/j.patcog.2023.109763 doi: 10.1016/j.patcog.2023.109763
|
| [36] |
T. W. Zhang, Y. K. Li, Global exponential stability of discrete-time almost automorphic Caputo-Fabrizio BAM fuzzy neural networks via exponential Euler technique, Knowl.-Based Syst., 246 (2022), 108675. https://doi.org/10.1016/j.knosys.2022.108675 doi: 10.1016/j.knosys.2022.108675
|
| [37] |
S. Rana, A. R. Bhowmick, S. Bhattacharya, Impact of prey refuge on a discrete time predator-prey system with Allee effect, Int. J. Bifurcat. Chaos, 24 (2014), 1450106. https://doi.org/10.1142/S0218127414501065 doi: 10.1142/S0218127414501065
|
| [38] |
R. Ma, Y. Z. Bai, F. Wang, Dynamical behavior analysis of a two-dimensional discrete predator-prey model with prey refuge and fear factor, J. Appl. Anal. Comput., 10 (2020), 1683–1697. https://doi.org/10.11948/20190426 doi: 10.11948/20190426
|
| [39] |
B. H. Hong, C. R. Zhang, Bifurcations and chaotic behavior of a predator-prey model with discrete time, AIMS Math., 8 (2023), 13390–13410. https://doi.org/10.3934/math.2023678 doi: 10.3934/math.2023678
|
| [40] |
Y. Huang, Z. Zhu, Z. Li, Modeling the Allee effect and fear effect in predator-prey system incorporating a prey refuge, Adv. Differ. Equ., 321 (2020), 1–13. https://doi.org/10.1186/s13662-020-02727-5 doi: 10.1186/s13662-020-02727-5
|
| [41] | A. C. Luo, Regularity and complexity in dynamical systems, New York: Springer, 2012. https://doi.org/10.1007/978-1-4614-1524-4 |
| [42] | G. Chen, X. Dong, From chaos to order: Perspectives, methodologies, and applications, Singapore: World Scientificr, 1998. https://doi.org/10.1142/3033 |
| [43] |
Q. Din, A. A. Elsadany, S. Ibrahim, Bifurcation analysis and chaos control in a second-order rational difference equation, Int. J. Nonlin. Sci. Num., 19 (2018), 53–68. https://doi.org/10.1515/ijnsns-2017-0077 doi: 10.1515/ijnsns-2017-0077
|
| [44] |
X. S. Luo, G. R. Chen, B. H. Wang, J. Q. Fang, Hybrid control of period-doubling bifurcation and chaos in discrete nonlinear dynamical systems, Chaos Soliton. Fract., 181 (2003), 775–783. https://doi.org/10.1016/s0960-0779(03)00028-6 doi: 10.1016/s0960-0779(03)00028-6
|
| [45] | T. S. Shores, Applied linear algebra and matrix analysis, Switzerland: Springer Cham, 2018. https://doi.org/10.1007/978-3-319-74748-4 |
| [46] | J. Carr, Applications of centre manifold theory, New York: Springer New York, 1982. https://doi.org/10.1007/978-1-4612-5929-9 |
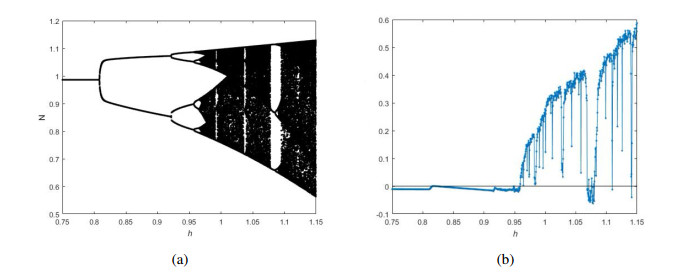
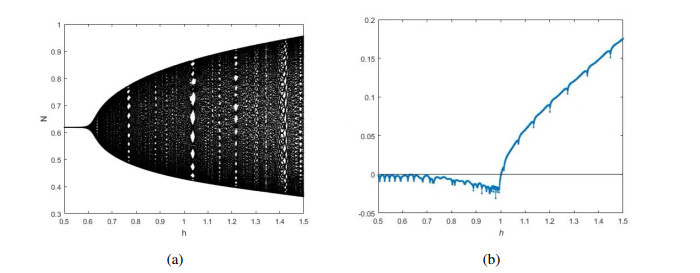
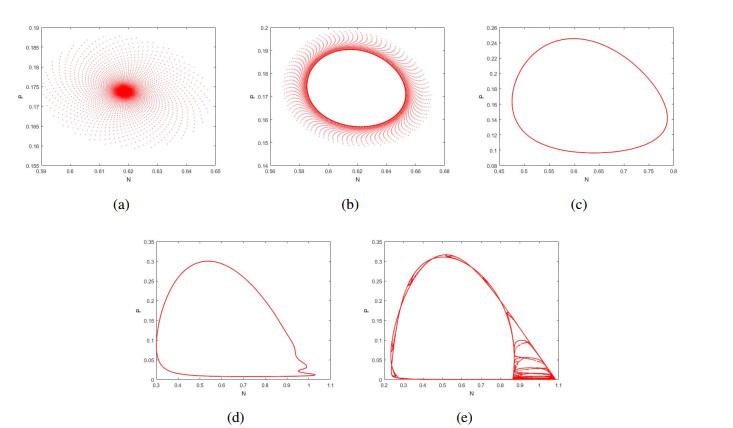
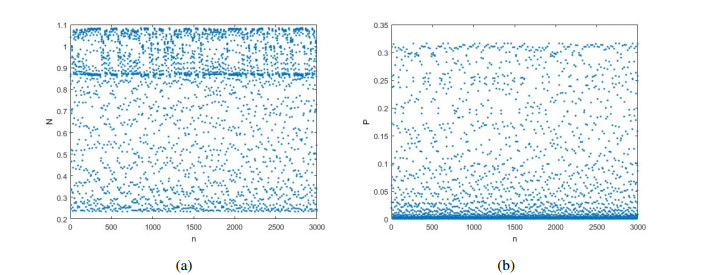
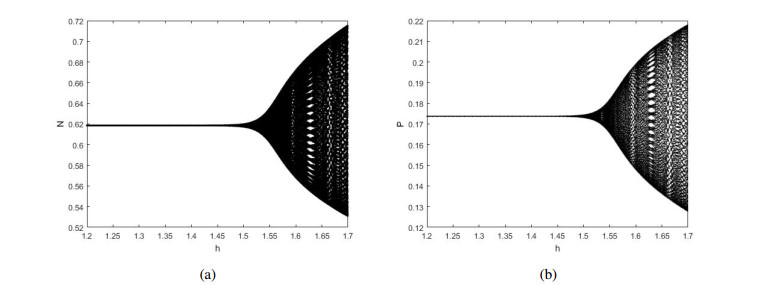
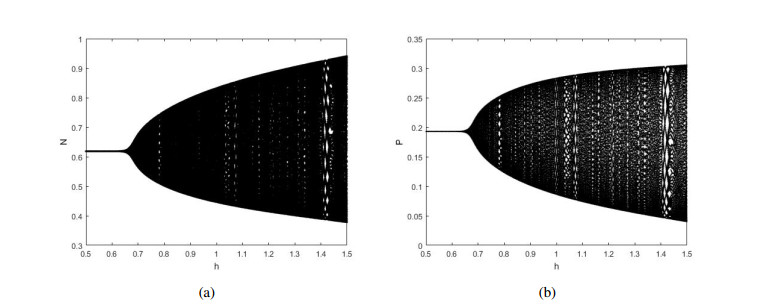
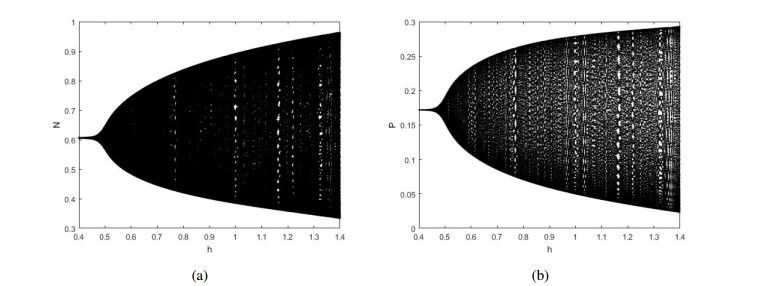
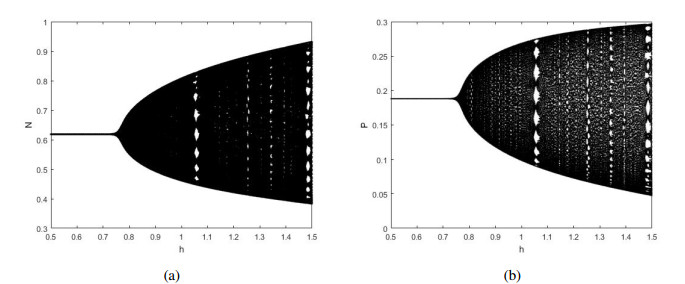
Figures(12)

Xiaoming Su, Jiahui Wang, Adiya Bao. Stability analysis and chaos control in a discrete predator-prey system with Allee effect, fear effect, and refuge[J]. AIMS Mathematics, 2024, 9(5): 13462-13491. doi: 10.3934/math.2024656







 DownLoad:
DownLoad: