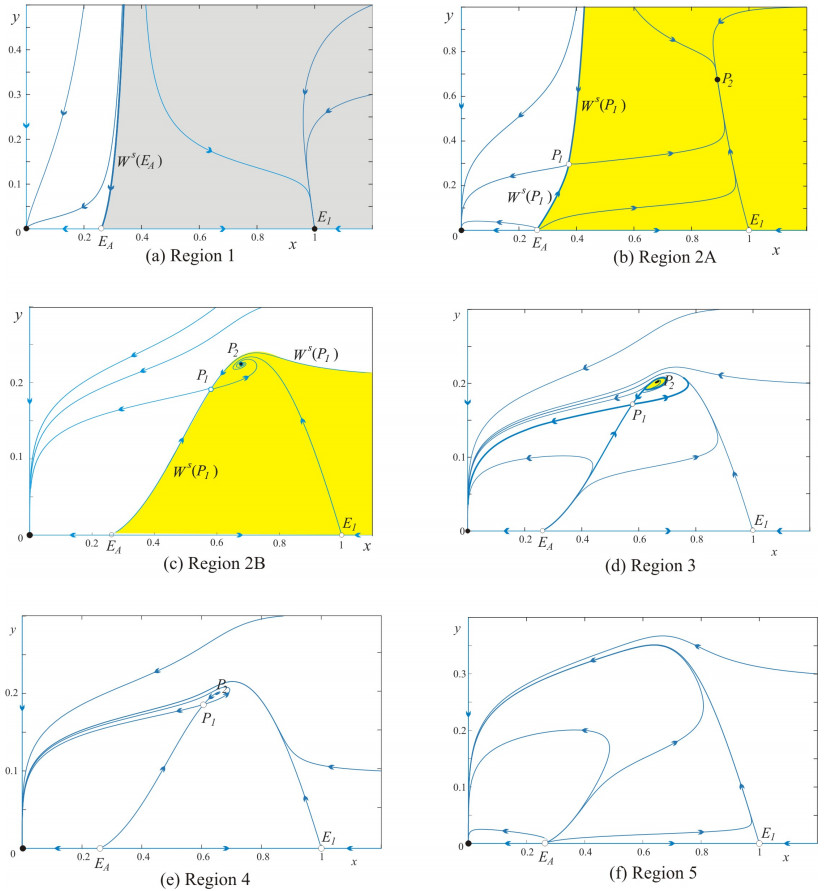
In this paper, we analyze a modified ratio-dependent predator-prey model with a strong Allee effect and linear prey refugee. The model exhibits rich dynamics with the existence of separatrices in the phase plane in-between basins of attraction associated with oscillation, coexistence, and extinction of the interacting populations. We prove that if the initial values are positive, all solutions are bounded and stay in the interior of the first quadrant. We show that the system undergoes several bifurcations such as transcritical, saddle-node, Hopf, and Bogdanov-Takens bifurcations. Consequently, a homoclinic bifurcation curve exists generating an unstable periodic orbit. Moreover, we find that the Bogdanov-Takens bifurcation acts as an organizing center for the scenario of surviving or extinction of both interacting species. Topologically different phase portraits with all possible trajectories and equilibria are depicted illustrating the behavior of the system.
Citation: Khairul Saleh. Basins of attraction in a modified ratio-dependent predator-prey model with prey refugee[J]. AIMS Mathematics, 2022, 7(8): 14875-14894. doi: 10.3934/math.2022816
In this paper, we analyze a modified ratio-dependent predator-prey model with a strong Allee effect and linear prey refugee. The model exhibits rich dynamics with the existence of separatrices in the phase plane in-between basins of attraction associated with oscillation, coexistence, and extinction of the interacting populations. We prove that if the initial values are positive, all solutions are bounded and stay in the interior of the first quadrant. We show that the system undergoes several bifurcations such as transcritical, saddle-node, Hopf, and Bogdanov-Takens bifurcations. Consequently, a homoclinic bifurcation curve exists generating an unstable periodic orbit. Moreover, we find that the Bogdanov-Takens bifurcation acts as an organizing center for the scenario of surviving or extinction of both interacting species. Topologically different phase portraits with all possible trajectories and equilibria are depicted illustrating the behavior of the system.

| [1] |
H. R. Akcakaya, R. Arditi, L. R. Ginzburg, Ratio-dependent prediction: An abstraction that works, Ecology, 76 (1995), 995–1004. https://doi.org/10.2307/1939362 doi: 10.2307/1939362

|
| [2] |
P. A. Abrams, L. R. Ginzburg, Coupling in predator-prey dynamics: Ratio-dependence, J. Theor. Biol., 139 (1989), 311–326. https://doi.org/10.1016/S0022-5193(89)80211-5 doi: 10.1016/S0022-5193(89)80211-5
|
| [3] |
P. Aguirre, J. D Flores, E. González-Olivares, Bifurcations and global dynamics in a predator-prey model with a strong Allee effect on the prey and ratio-dependent functional response, Nonlinear Anal.: Real Word Appl., 16 (2014), 235–249. https://doi.org/10.1016/j.nonrwa.2013.10.002 doi: 10.1016/j.nonrwa.2013.10.002
|
| [4] |
V. Ajraldi, M. Pittavino, E. Venturino, Modeling herd behavior in population systems, Nonlinear Anal.: Real Word Appl., 12 (2011), 2319–2338. https://doi.org/10.1016/j.nonrwa.2011.02.002 doi: 10.1016/j.nonrwa.2011.02.002
|
| [5] | W. C. Allee, Animal aggregations: A study in general sociology, USA: University of Chicago press, 1931. https://doi.org/10.5962/bhl.title.7313 |
| [6] | J. Bascompte, Extinction thresholds: Insights from simple models, Ann. Zool. Fenn., 40 (2003), 99–114. |
| [7] |
S. Bentout, S. Djilali, A. Atangana, Bifurcation analysis of an age‐structured prey-predator model with infection developed in prey, Math. Methods Appl. Sci., 45 (2022), 1189–1208. https://doi.org/10.1002/mma.7846 doi: 10.1002/mma.7846
|
| [8] |
F. Berezovskaya, G. Karev, R. Arditi, Parametric analysis of the ratio-dependent predator-prey model, J. Math. Biol., 43 (2001), 221–246. https://doi.org/10.1007/s002850000078 doi: 10.1007/s002850000078
|
| [9] |
L. Berec, E. Angulo, F. Courchamp, Multiple Allee effects and population management, Trends Ecol. Evol., 22 (2007), 185–191. https://doi.org/10.1016/j.tree.2006.12.002 doi: 10.1016/j.tree.2006.12.002
|
| [10] |
H. W. Broer, K. Saleh, V. Naudot, R. Roussarie, Dynamics of a predator-prey model with non-monotonic response function, Discrete Cont. Dyn. Syst., 18 (2007), 221–251. https://doi.org/10.3934/dcds.2007.18.221 doi: 10.3934/dcds.2007.18.221
|
| [11] |
A. A. Berryman, The origins and evolutions of predator-prey theory, Ecology, 73 (1992), 1530–1535. https://doi.org/10.2307/1940005 doi: 10.2307/1940005
|
| [12] |
L. Chen, F. Chen, L. Chen, Qualitative analysis of a predator-prey model with Holling type II functional response incorporating a constant prey refuge, Nonlinear Anal: Real Word Appl., 11 (2010), 246–252. https://doi.org/10.1016/j.nonrwa.2008.10.056 doi: 10.1016/j.nonrwa.2008.10.056
|
| [13] |
D. L. DeAngelis, R. A. Goldstein, R. V. O'Neill, A model for trophic interactions, Ecology, 56 (1975), 881–892. https://doi.org/10.2307/1936298 doi: 10.2307/1936298
|
| [14] |
B. Dennis, Allee effects: Population growth, critical density, and change of extinction, Nat. Resour. Model., 3 (1989), 481–538. https://doi.org/10.1111/j.1939-7445.1989.tb00119.x doi: 10.1111/j.1939-7445.1989.tb00119.x
|
| [15] |
A. Dhooge, W. Govaerts, Y. Kuznetsov, MATCONT: A MATLAB package for numerical bifurcation analysis of ODEs, ACM Trans. Math. Sofware, 29 (2003), 141–164. https://doi.org/10.1145/779359.779362 doi: 10.1145/779359.779362
|
| [16] |
S. Djilali, S. Bentout, Pattern formations of a delayed diffusive predator-prey model with predator harvesting and prey social behavior, Math. Methods Appl. Sci., 44 (2021), 9128–9142. https://doi.org/10.1002/mma.7340 doi: 10.1002/mma.7340
|
| [17] |
M. Fan, P. Wu, Z. Feng, R. K. Swihar, Dynamics of predator-prey metapopulations with Allee effects, Bull. Math. Biol., 78 (2016), 1727–1748. https://doi.org/10.1007/s11538-016-0197-6 doi: 10.1007/s11538-016-0197-6
|
| [18] |
J. D. Flores, E. González-Olivares, Dynamics of a predator-prey model with Allee effect on prey and ratio-dependent functional response, Ecol. Complex., 18 (2014), 59–66. https://doi.org/10.1016/j.ecocom.2014.02.005 doi: 10.1016/j.ecocom.2014.02.005
|
| [19] | H. I. Freedman, Deterministic mathematical model in population ecology, Marcel Dekker, New York, 1980. |
| [20] |
J. Gascoigne, R. N. Lipcius, Allee effects driven by predation, J. Appl. Ecol., 41 (2004), 801–810. https://doi.org/10.1111/j.0021-8901.2004.00944.x doi: 10.1111/j.0021-8901.2004.00944.x
|
| [21] |
S. Gregory, F. Courchamp, Safety in numbers: Extinction arising from predator-driven Allee effects, J. Anim. Ecol., 79 (2010), 511–514. https://doi.org/10.1111/j.1365-2656.2010.01676.x doi: 10.1111/j.1365-2656.2010.01676.x
|
| [22] |
Y. Gao, B. Li, Dynamics of a ratio-dependent predator-prey system with strong Allee effect, Discrete Cont. Dyn. Syst., 18 (2013), 2283–2313. https://doi.org/10.3934/dcdsb.2013.18.2283 doi: 10.3934/dcdsb.2013.18.2283
|
| [23] |
L. R. Ginzburg, H. R. Akcakaya, Consequences of ratio-dependent predation for steady-state properties of ecosystems, Ecology, 73 (1992), 1536–1543. https://doi.org/10.2307/1940006 doi: 10.2307/1940006
|
| [24] |
E. González-Olivars, R. Ramos-Jiliberto, Dynamics consequences of prey refuges in a simple model system: More prey, few predators and enhanced stability, Ecol. Model., 166 (2003), 135–146. https://doi.org/10.1016/S0304-3800(03)00131-5 doi: 10.1016/S0304-3800(03)00131-5
|
| [25] |
A. P. Gutierrez, Physiological basis of ratio-dependent predator-prey theory: A metabolic pool model of Nicholson's blowflies as an example, Ecology, 73 (1992), 1552–1563. https://doi.org/10.2307/1940008 doi: 10.2307/1940008
|
| [26] |
M. Haque, Ratio-dependent predator-prey models of interacting populations, Bull. Math. Biol., 71 (2009), 430–452. https://doi.org/10.1007/s11538-008-9368-4 doi: 10.1007/s11538-008-9368-4
|
| [27] |
M. Haque, M. S. Rahman, E. Venturino, B. L. Li, Effect of a functional response-dependent prey refuge in a predator-prey model, Ecol. Complex., 20 (2014), 248–256. https://doi.org/10.1016/j.ecocom.2014.04.001 doi: 10.1016/j.ecocom.2014.04.001
|
| [28] |
C. S. Holling, The functional response of predators to prey density and its role in mimicry and population regulation, Mem. Entomol. Soc. Can., 97 (1965), 3–60. https://doi.org/10.4039/entm9745fv doi: 10.4039/entm9745fv
|
| [29] |
S. B. Hsu, T. W. Hwang, Y. Kuang, Global analysis of the Michaelis-Menten type ratio-dependent predator-prey system, J. Math. Biol., 42 (2001), 489–506. https://doi.org/10.1007/s002850100079 doi: 10.1007/s002850100079
|
| [30] |
Y. Huang, F. Chen, L. Zhong, Stability analysis of a prey-predator model with Holling type III response function incorporating a prey refuge, Appl. Math. Comput., 182 (2006), 672–683. https://doi.org/10.1016/j.amc.2006.04.030 doi: 10.1016/j.amc.2006.04.030
|
| [31] |
T. K. Kar, Stability analysis of a prey-predator model incorporating a prey refuge, Commun. Nonlinear Sci. Numer. Simul., 10 (2005), 681–691. https://doi.org/10.1016/j.cnsns.2003.08.006 doi: 10.1016/j.cnsns.2003.08.006
|
| [32] |
Y. Kuang, E. Beretta, Global qualitative analysis of a ratio-dependent predator-prey system, J. Math. Biol., 36 (1998), 389–406. https://doi.org/10.1007/s002850050105 doi: 10.1007/s002850050105
|
| [33] |
V. Krivan, Effects of optimal antipredator behavior of prey on predator-prey dynamics: The role of refuges, Theor. Popul. Biol., 53 (1998), 131–142. https://doi.org/10.1006/tpbi.1998.1351 doi: 10.1006/tpbi.1998.1351
|
| [34] | Y. A. Kuznetsov, Elements of applied bifurcation theory, Springer, New York, 1998. |
| [35] | A. J. Lotka, Elements of physical biology, Williams and Wilkins, Baltimore MD, 1925. |
| [36] |
Z. Ma, S. Wang, W. Li, Z. Li, The effect of prey refuge in a patchy predator-prey system, Math. Biosci., 243 (2013), 126–130. https://doi.org/10.1016/j.mbs.2013.02.011 doi: 10.1016/j.mbs.2013.02.011
|
| [37] | J. D. Murray, Mathematical biology (Biomathematics, Vol. 19), New York: Springer Verlag, 1993. |
| [38] |
J. N. McNair, The effects of refuges on predator-prey interactions: A reconsideration, Theor. Popul. Biol., 29 (1986), 38–63. https://doi.org/10.1016/0040-5809(86)90004-3 doi: 10.1016/0040-5809(86)90004-3
|
| [39] |
A. Morozov, S. Petrovoskii, B. L. Li, Spatiotemporal complexity of patchy invasion in a predator-prey system with Allee effect, J. Theor. Biol., 238 (2006), 18–35. https://doi.org/10.1016/j.jtbi.2005.05.021 doi: 10.1016/j.jtbi.2005.05.021
|
| [40] | L. Perko, Differential equations and dynamical systems, New York: Springer, 2001. https://doi.org/10.1007/978-1-4613-0003-8 |
| [41] |
G. D. Ruxton, Short term refuge use and stability of predator-prey models, Theor. Popul. Biol., 47 (1995), 1–17. https://doi.org/10.1006/tpbi.1995.1001 doi: 10.1006/tpbi.1995.1001
|
| [42] |
K. Saleh, Dynamics of a predator-prey model with Allee effect and prey group defense, AIP Conf. Proc., 1643 (2015), 655–661. https://doi.org/10.1063/1.4907508 doi: 10.1063/1.4907508
|
| [43] |
P. A. Stephens, W. J. Sutherland, Consequences of the Allee effect for behaviour, ecology and conservation, Trends Ecol. Evol., 14 (1999), 401–405. https://doi.org/10.1016/s0169-5347(99)01684-5 doi: 10.1016/s0169-5347(99)01684-5
|
| [44] | V. Volterra, Variazioni e fluttuaziono del numero di individui in specie animali conviventi, Memoria della Reale Accad. Nazionale dei Lincei, 2 (1926), 31–113. |
| [45] |
G. S. W. Wolkowicz, Bifurcation analysis of a predator-prey system involving group defense, SIAM J. Appl. Math., 48 (1988), 592–606. https://doi.org/10.1137/0148033 doi: 10.1137/0148033
|
| [46] |
D. Xiao, W. Li, M. Han, Dynamics in a ratio-dependent predator-prey model with predator harvesting, J. Math. Anal. Appl., 324 (2006), 14–29. https://doi.org/10.1016/j.jmaa.2005.11.048 doi: 10.1016/j.jmaa.2005.11.048
|
| [47] |
D. Xiao, S. Ruan, Codimension two bifurcations in a predator-prey system with group defense, Int. J. Bifurcat. Chaos, 11 (2001), 2123–2131. https://doi.org/10.1142/S021812740100336X doi: 10.1142/S021812740100336X
|
| [48] |
D. Xiao, S. Ruan, Global dynamics of a ratio-dependent predator-prey system, J. Math. Biol., 43 (2001), 268–290. https://doi.org/10.1007/s002850100097 doi: 10.1007/s002850100097
|




Figures(5) / Tables(1)

Khairul Saleh. Basins of attraction in a modified ratio-dependent predator-prey model with prey refugee[J]. AIMS Mathematics, 2022, 7(8): 14875-14894. doi: 10.3934/math.2022816







 DownLoad:
DownLoad: