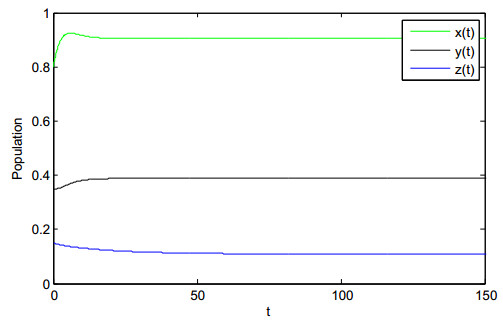
In this paper, we consider a stochastic two predator-one prey system consisting of prey, intermediate predator and top predator with Lévy jumps. Here we consider Ratio-dependent function response between intermediate predator and top predator and other function responses are assumed to be linear. Firstly, we prove that the existence and boundedness of pth moment of the positive solution. Then under some assumptions, we establish sufficient criteria for the extinction of the system. The results reveal an important property that the Lévy jumps are unfavorable for the existence of species. Furthermore, we establish sufficient condition for the asymptotically stable in distribution under certain conditions. Finally, some numerical simulations are introduced to demonstrate the theoretical results.
Citation: Xuegui Zhang, Yuanfu Shao. Analysis of a stochastic predator-prey system with mixed functional responses and Lévy jumps[J]. AIMS Mathematics, 2021, 6(5): 4404-4427. doi: 10.3934/math.2021261
In this paper, we consider a stochastic two predator-one prey system consisting of prey, intermediate predator and top predator with Lévy jumps. Here we consider Ratio-dependent function response between intermediate predator and top predator and other function responses are assumed to be linear. Firstly, we prove that the existence and boundedness of pth moment of the positive solution. Then under some assumptions, we establish sufficient criteria for the extinction of the system. The results reveal an important property that the Lévy jumps are unfavorable for the existence of species. Furthermore, we establish sufficient condition for the asymptotically stable in distribution under certain conditions. Finally, some numerical simulations are introduced to demonstrate the theoretical results.

| [1] |
N. Ali, S. Chakravarty, Stability analysis of a food chain model consisting of two competitive preys and one predator, Nonlinear Dyn., 82 (2015), 1303–1316. doi: 10.1007/s11071-015-2239-2

|
| [2] | M. Elettreby, Two-prey one-predator model, Chaos Solitons Fractals, 39 (2009), 2018–2027. |
| [3] |
S. Ahmad, I. M. Stamova, Almost necessary and sufficient conditions for survival of species, Nonlinear Anal.: RWA, 5 (2004), 219–229. doi: 10.1016/S1468-1218(03)00037-3
|
| [4] |
M. Farkas, H. Freedman, Stability conditions for two predator one prey system, Acta Appl. Math., 14 (1989), 3–10. doi: 10.1007/BF00046669
|
| [5] |
A. Hastings, T. Powell, Chaos in a three-species food chain, Ecology, 72 (1991), 896–903. doi: 10.2307/1940591
|
| [6] |
A. L. Koch, Competitive coexistence of two predators utilizing the same prey under rconstant enviromental conditions, J. Theor. Biol., 44 (1974), 387–395. doi: 10.1016/0022-5193(74)90169-6
|
| [7] |
R. M. May, W. J. Leonard, Nonlinear aspects of competition between three species, SIAM J. Appl. Math., 29 (1975), 243–253. doi: 10.1137/0129022
|
| [8] |
H. I. Freedman, P. Waltman, Persistence in models of three interacting predator-prey populations, Math. Biosci., 68 (1984), 213–231. doi: 10.1016/0025-5564(84)90032-4
|
| [9] |
S. Hsu, S. Ruan, T. Yang, Analysis of three species Lotka-Volterra food web models with omnivory, J. Math. Anal. Appl., 426 (2015), 659–687. doi: 10.1016/j.jmaa.2015.01.035
|
| [10] |
D. Sen, S. Ghorai, M. Banerjee, Complex dynamics of a three species prey-predator model with intraguild predation, Ecol. Complexity, 34 (2018), 9–22. doi: 10.1016/j.ecocom.2018.02.002
|
| [11] | X. Mao, Stochastic differential equations and applications, Horwood Publishing, Chichester, 1997. |
| [12] |
M. Wang, W. Li, Stability of random impulsive coupled systems on networks with Markovian switching, Stoch. Anal. Appl., 37 (2019), 1107–1132. doi: 10.1080/07362994.2019.1643247
|
| [13] |
H. Zhou, Y. Zhang, W. Li, Synchronization of stochastic Lévy noise systems on a multi-weights network and its applications of chua's circuits, IEEE Trans. Circuits Syst. I, 66 (2019), 2709–2722. doi: 10.1109/TCSI.2019.2899375
|
| [14] |
H. Zhou, W. Li, Synchronisation of stochastic-coupled intermittent control systems with delays and Lévy noise on networks without strong connectedness, IET Control Theory Appl., 13 (2019), 36–49. doi: 10.1049/iet-cta.2018.5187
|
| [15] |
Q. Liu, Q. Chen, Y. Hu, Analysis of a stochastic mutualism model, Commun. Nonlinear Sci. Numer. Simul., 29 (2015), 188–197. doi: 10.1016/j.cnsns.2015.05.010
|
| [16] |
P. Wang, B. Zhang, H. Su, Stabilization of stochastic uncertain complex-valued delayed networks via aperiodically intermittent nonlinear control, IEEE Trans. Syst. Man Cybern.: Syst., 49 (2019), 649–662. doi: 10.1109/TSMC.2018.2818129
|
| [17] | R. M. May, Stability and complexity in model ecosystems, Princeton University Press, NJ, 2001. |
| [18] |
X. H. Zhang, W. X. Li, M. Liu, K. Wang, Dynamics of a stochastic Holling II one-predator two-prey system with jumps, Phys. A, 421 (2015), 571–582. doi: 10.1016/j.physa.2014.11.060
|
| [19] |
X. Mao, G. Marion, E. Renshaw, Environmental Brownian noise suppresses explosions in population dynamics, Stoch. Process. Appl., 97 (2002), 95–110. doi: 10.1016/S0304-4149(01)00126-0
|
| [20] |
Q. Liu, D. Jiang, T. Hayat, A. Alsaedi, Dynamics of a stochastic predator-prey model with stage structure for predator and holling type II functional response, J. Nonlinear Sci., 28 (2018), 1151–1187. doi: 10.1007/s00332-018-9444-3
|
| [21] |
J. Bao, X. Mao, G. Yin, C. Yuan, Competitive Lotka-Volterra population dynamics with jumps, Nonlinear Anal., 74 (2011), 6601–6616. doi: 10.1016/j.na.2011.06.043
|
| [22] |
J. Bao, C. Yuan, Stochastic population dynamics driven by Lévy noise, J. Math. Anal. Appl., 391 (2012), 363–375. doi: 10.1016/j.jmaa.2012.02.043
|
| [23] |
Q. Liu, Asymptotic properties of a stochastic n-species Gilpin-Ayala competitive model with Lévy jumps and Markovian switching, Commun. Nonlinear Sci Numer. Simul., 26 (2015), 1–10. doi: 10.1016/j.cnsns.2015.01.007
|
| [24] | X. Mao, C. Yuan, Stochastic differential equations with Markovian switching, Imperial College Press, London, 2006. |
| [25] |
M. Liu, K. Wang, Stochastic Lotka-Volterra systems with Lévy noise, J. Math. Anal. Appl., 410 (2014), 750–763. doi: 10.1016/j.jmaa.2013.07.078
|
| [26] |
R. Liu, G. Liu, Analysis on stochastic food-web model with intraguild predation and mixed functional responses, Phys. A, 531 (2019), 121782. doi: 10.1016/j.physa.2019.121782
|
| [27] |
D. Sen, S. Ghorai, M. Banerjee, Complex dynamics of a three species prey-predator model with intraguild predation, Ecol. Complex, 34 (2018), 9–22. doi: 10.1016/j.ecocom.2018.02.002
|
| [28] |
X. Mao, Stochastic versions of the Lassalle theorem, J. Differ. Equations, 153 (1999), 175–195. doi: 10.1006/jdeq.1998.3552
|
| [29] | I. Karatzas, S. E. Shreve, Brownian motion and stochastic calculus, Springer-Verlag, Berlin, New York, 1991. |
| [30] | A. Friedman, Stochastic differential equations and their applications, Academic Press, New York, 1976. |
| [31] | D. Applebaum, Lévy processes and stochastic calculus, 2 Eds., Cam-bridge Unversity Press, 2009. |
| [32] | D. J. Higham, An algorithmic introduction to numerical simulation of stochastic differential equations, SIAM Rev., 43 (2001) 525–546. |
| [33] |
Q. Liu, D. Jiang, T. Hayat, A. Alsaedi, Dynamics of a stochastic predator-prey model with distributed delay and Markovian switching, Phys. A, 527 (2019), 121264. doi: 10.1016/j.physa.2019.121264
|
| [34] |
Z. Wang, M. Deng, M. Liu, Stationary distribution of a stochastic ratio-dependent predator-prey system with regime-switching, Chaos, Solitons Fractals, 142 (2021), 110462. doi: 10.1016/j.chaos.2020.110462
|
| [35] | M. Liu, C. Bai, Optimal harvesting of a stochastic mutualism model with regime-switching, Appl. Math. Comput., 373 (2020), 125040. |
| [36] |
C. Lu, Dynamics of a stochastic Markovian switching predator-prey model with infinite memory and general Lévy jumps, Math. Comput. Simul., 181 (2021), 316–332. doi: 10.1016/j.matcom.2020.10.002
|
| [37] |
H. Wang, D. Jiang, T. Hayat, A. Alsaedic, B. Ahmad, Stationary distribution of stochastic NP ecological model under regime switching, Phys. A, 549 (2020), 124064. doi: 10.1016/j.physa.2019.124064
|
| [38] | C. Lu, X. Ding, Periodic solutions and stationary distribution for a stochastic predator-prey system with impulsive perturbations, Appl. Math. Comput., 350 (2019), 313–322. |
| [39] |
B. Han, D. Jiang, T. Hayat, A. Alsaedic, B. Ahmad, Stationary distribution and extinction of a stochastic staged progression AIDS model with staged treatment and second-order perturbation, Chaos, Solitons Fractals, 140 (2020), 110238. doi: 10.1016/j.chaos.2020.110238
|



Figures(4)

Xuegui Zhang, Yuanfu Shao. Analysis of a stochastic predator-prey system with mixed functional responses and Lévy jumps[J]. AIMS Mathematics, 2021, 6(5): 4404-4427. doi: 10.3934/math.2021261







 DownLoad:
DownLoad: