This paper studies the mixed passive and $ H_{\infty} $ performance for uncertain neural networks with interval discrete and distributed time-varying delays via feedback control. The interval discrete and distributed time-varying delay functions are not assumed to be differentiable. The improved criteria of exponential stability with a mixed passive and $ H_{\infty} $ performance are obtained for the uncertain neural networks by constructing a Lyapunov-Krasovskii functional (LKF) comprising single, double, triple, and quadruple integral terms and using a feedback controller. Furthermore, integral inequalities and convex combination technique are applied to achieve the less conservative results for a special case of neural networks. By using the Matlab LMI toolbox, the derived new exponential stability with a mixed passive and $ H_{\infty} $ performance criteria is performed in terms of linear matrix inequalities (LMIs) that cover $ H_{\infty} $, and passive performance by setting parameters in the general performance index. Numerical examples are shown to demonstrate the benefits and effectiveness of the derived theoretical results. The method given in this paper is less conservative and more general than the others.
Citation: Sunisa Luemsai, Thongchai Botmart, Wajaree Weera, Suphachai Charoensin. Improved results on mixed passive and $ H_{\infty} $ performance for uncertain neural networks with mixed interval time-varying delays via feedback control[J]. AIMS Mathematics, 2021, 6(3): 2653-2679. doi: 10.3934/math.2021161
This paper studies the mixed passive and $ H_{\infty} $ performance for uncertain neural networks with interval discrete and distributed time-varying delays via feedback control. The interval discrete and distributed time-varying delay functions are not assumed to be differentiable. The improved criteria of exponential stability with a mixed passive and $ H_{\infty} $ performance are obtained for the uncertain neural networks by constructing a Lyapunov-Krasovskii functional (LKF) comprising single, double, triple, and quadruple integral terms and using a feedback controller. Furthermore, integral inequalities and convex combination technique are applied to achieve the less conservative results for a special case of neural networks. By using the Matlab LMI toolbox, the derived new exponential stability with a mixed passive and $ H_{\infty} $ performance criteria is performed in terms of linear matrix inequalities (LMIs) that cover $ H_{\infty} $, and passive performance by setting parameters in the general performance index. Numerical examples are shown to demonstrate the benefits and effectiveness of the derived theoretical results. The method given in this paper is less conservative and more general than the others.

| [1] |
L. O. Chua, L. Yang, Cellular neural networks: applications, IEEE Trans. Circuits Syst., 35 (1988), 1273–1290. doi: 10.1109/31.7601

|
| [2] | M. A. Cohen, S. Grossberg, Absolute stability of global pattern formation and parallel memory storage by competitive neural networks, IEEE Trans. Syst. Man Cybern., 13 (1983), 815–826. |
| [3] | S. Haykin, Neural Networks, New Jersey: Englewood Cliffs, 1994. |
| [4] |
H. Wang, Y. Yu, G. Wen, Stability analysis of fractional-order Hopfield neural networks with time delays, Neural Networks, 55 (2014), 98–109. doi: 10.1016/j.neunet.2014.03.012
|
| [5] | H. Zhang, Z. Wang, New delay-dependent criterion for the stability of recurrent neural networks with time-varying delay, Sci. China Series F, 52 (2009), 942–948. |
| [6] |
L. Wang, X. Zou, Harmless delays in Cohen-Grossberg neural networks, Phys. D, 170 (2002), 162–173. doi: 10.1016/S0167-2789(02)00544-4
|
| [7] | A. Farnam, R. M. Esfanjani, A. Ahmadi, Delay-dependent criterion for exponential stability analysis of neural networks with time-varying delays, IFAC-PapersOnLine, 49 (2016), 130–135. |
| [8] |
H. B. Zeng, Y. He, P. Shi, M. Wu, S. P. Xiao, Dissipativity analysis of neural networks with time-varying delays, Neurocomputing, 168 (2015), 741–746. doi: 10.1016/j.neucom.2015.05.050
|
| [9] |
Y. Liu, Z. Wang, X. Liu, Global exponential stability of generalized recurrent neural networks with discrete and distributed delays, Neural Networks, 19 (2006), 667–675. doi: 10.1016/j.neunet.2005.03.015
|
| [10] | H. P. Kriegel, M. Pfeifle, Density-based clustering of uncertain data, In: Proceedings of the ACM SIGKDD International Conference on Knowledge Discovery and Data Mining, 2005,672–677. |
| [11] | G. Cormode, A. McGregor, Approximation algorithms for clustering uncertain data, In: Proceedings of the ACM SIGACT-SIGMOD-SIGART Symposium on Principles of Database Systems, 2008,191–199. |
| [12] | C. C. Aggarwal, P. Yu, A framework for clustering uncertain data streams, In: Proceedings-International Conference on Data Engineering, 2008,150–159. |
| [13] | K. Subramanian, P. Muthukumar, S. Lakshmanan, Robust stabilization of uncertain neural networks with additive time-varying delays, IFAC-PapersOnLine, 49 (2016), 154–159. |
| [14] |
H. B. Zeng, J. H. Park, H. Shen, Robust passivity analysis of neural networks with discrete and distributed delays, Neurocomputing, 149 (2015), 1092–1097. doi: 10.1016/j.neucom.2014.07.024
|
| [15] |
L. Wu, W. X. Zheng, Passivity-based sliding mode control of uncertain singular time-delay systems, Automatica, 45 (2009), 2120–2127. doi: 10.1016/j.automatica.2009.05.014
|
| [16] |
G. Calcev, R. Gorez, M. D. Neyer, Passivity approach to fuzzy control systems, Automatica, 34 (1998), 339–344. doi: 10.1016/S0005-1098(97)00202-1
|
| [17] |
H. Gao, T. Chen, T. Chai, Passivity and passification for networked control systems, SIAM J. Control Optim., 46 (2007), 1299–1322. doi: 10.1137/060655110
|
| [18] |
L. Xie, M. Fu, H. Li, Passivity analysis and passification for uncertain signal processing systems, IEEE Trans. Signal Process., 46 (1998), 2394–2403. doi: 10.1109/78.709527
|
| [19] | H. Li, J. Lam, K. C. Cheung, Passivity criteria for continuous-time neural networks with mixed time-varying delays, Appl. Math. Comput., 218 (2012), 11062–11074. |
| [20] |
M. V. Thuan, H. Trinh, L. V. Hien, New inequality-based approach to passivity analysis of neural networks with interval time-varying delay, Neurocomputing, 194 (2016), 301–307. doi: 10.1016/j.neucom.2016.02.051
|
| [21] | S. Xu, W. X. Zheng, Y. Zou, Passivity analysis of neural networks with time-varying delays, IEEE T. Circuits II, 56 (2009), 325–329. |
| [22] |
N. Yotha, T. Botmart, K. Mukdasai, W. Weera, Improved delay-dependent approach to passivity analysis for uncertain neural networks with discrete interval and distributed time-varying delays, Vietnam J. Math., 45 (2017), 721–736. doi: 10.1007/s10013-017-0243-1
|
| [23] |
Y. Du, X. Liu, S. Zhong, Robust reliable $H_\infty$ control for neural networks with mixed time delays, Chaos Soliton. Fract., 91 (2016), 1–8. doi: 10.1016/j.chaos.2016.04.009
|
| [24] |
M. Syed Ali, R. Saravanakumar, S. Arik, Novel $H_{\infty}$ state estimation of static neural networks with interval time-varying delays via augmented Lyapunov-Krasovskii functional, Neurocomputing, 171 (2016), 949–954. doi: 10.1016/j.neucom.2015.07.038
|
| [25] |
K. Mathiyalagan, J. H. Park, R. Sakthivel, S. M. Anthoni, Robust mixed $H_\infty$ and passive filtering for networked markov jump systems with impulses, Signal Process., 101 (2014), 162–173. doi: 10.1016/j.sigpro.2014.02.007
|
| [26] |
M. Meisami-Azad, J. Mohammadpour, K. M. Grigoriadis, Dissipative analysis and control of state-space symmetric systems, Automatica, 45 (2009), 1574–1579. doi: 10.1016/j.automatica.2009.02.015
|
| [27] | L. Su, H. Shen, Mixed $H_{\infty}$/passive synchronization for complex dynamical networks with sampled-data control, Appl. Math. Comput., 259 (2015), 931–942. |
| [28] |
J. Wang, L. Su, H. Shen, Z. G. Wu, J. H. Park, Mixed $H_\infty$/passive sampled-data synchronization control of complex dynamical networks with distributed coupling delay, J. Franklin Inst., 354 (2017), 1302–1320. doi: 10.1016/j.jfranklin.2016.11.035
|
| [29] |
R. Sakthivel, R. Anbuvithya, K. Mathiyalagan, P. Prakash, Combined $H_{\infty}$ and passivity state estimation of memristive neural networks with random gain fluctuations, Neurocomputing, 168 (2015), 1111–1120. doi: 10.1016/j.neucom.2015.05.012
|
| [30] |
J. Qiu, H. Yang, J. Zhang, Z. Gao, New robust stability criteria for uncertain neural networks with interval time-varying delays, Chaos Soliton. Fract., 39 (2009), 579–585. doi: 10.1016/j.chaos.2007.01.087
|
| [31] |
J. Sun, G. P. Liu, J. Chen, D. Rees, Improved stability criteria for neural networks with time-varying delay, Phys. Lett. A, 373 (2009), 342–348. doi: 10.1016/j.physleta.2008.11.048
|
| [32] |
J. Tian, X. Xie, New asymptotic stability criteria for neural networks with time-varying delay, Phys. Lett. A, 374 (2010), 938–943. doi: 10.1016/j.physleta.2009.12.020
|
| [33] |
A. Farnam, R. M. Esfanjani, Improved linear matrix inequality approach to stability analysis of linear systems with interval time-varying delays, J. Comput. Appl. Math., 294 (2016), 49–56. doi: 10.1016/j.cam.2015.07.031
|
| [34] |
J. An, Z. Li, X. Wang, A novel approach to delay-fractional dependent stability criterion for linear systems with interval delay, ISA Trans., 53 (2014), 210–219. doi: 10.1016/j.isatra.2013.11.020
|
| [35] | A. Seuret, F. Gouaisbaut, Jensen's and Wirtinger's inequalities for time-delay systems, In: Proceedings of the 11th IFAC Workshop on Time-Delay Systems, 2013,343–348. |
| [36] |
Z. Wang, Y. Liu, K. Fraser, X. Liu, Stochastic stability of uncertain Hopfield neural networks with discrete and distributed delays, Phys. Lett. A, 354 (2006), 288–297. doi: 10.1016/j.physleta.2006.01.061
|
| [37] | B. Boyd, L. E. Ghoui, E. Feron, V. Balakrishnan, Linear matrix inequalities in system and control theory, Philadephia: SIAM, 1994. |
| [38] |
Y. He, G. P. Liu, D. Rees, M. Wu, Stability analysis for neural networks with time-varying interval delay, IEEE Trans. Neural Netw., 18 (2007), 1850–1854. doi: 10.1109/TNN.2007.903147
|
| [39] | G. Balas, R. Chaing, A. Packard, M. Safovov, Robust control toolbox user's guide, Natick: The MathWorks, 2010. |
| [40] |
P. Niamsup, T. Botmart, W. Weera, Modified function projective synchronization of complex dynamical networks with mixed time-varying and asymmetric coupling delays via new hybrid pinning adaptive control, Adv. Differ. Equ., 2017 (2017), 1–31. doi: 10.1186/s13662-016-1057-2
|
| [41] |
H. Shu, Q. Song, Y. Liu, Z. Zhao, F. E. Alsaadi, Global $\mu$-stability of quaternion-valued neural networks with non-differentiable time-varying delays, Neurocomputing, 247 (2017), 202–212. doi: 10.1016/j.neucom.2017.03.052
|
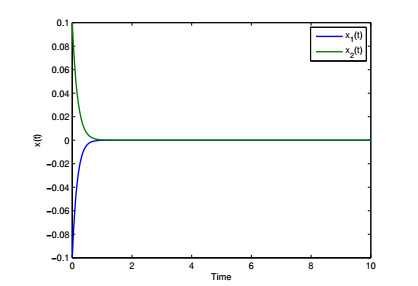
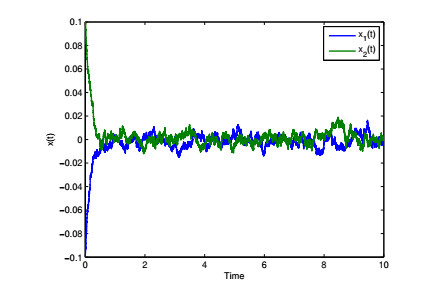
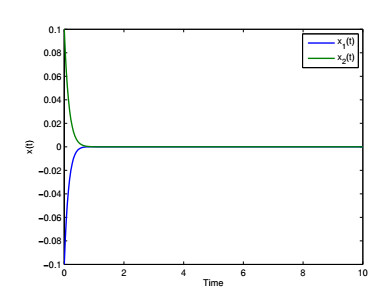
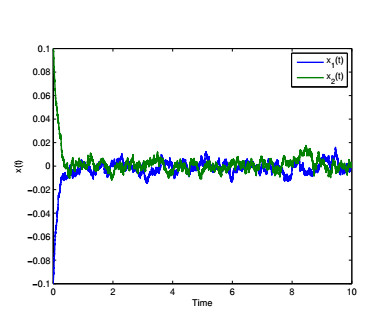
Figures(4) / Tables(6)

Sunisa Luemsai, Thongchai Botmart, Wajaree Weera, Suphachai Charoensin. Improved results on mixed passive and $ H_{\infty} $ performance for uncertain neural networks with mixed interval time-varying delays via feedback control[J]. AIMS Mathematics, 2021, 6(3): 2653-2679. doi: 10.3934/math.2021161







 DownLoad:
DownLoad: