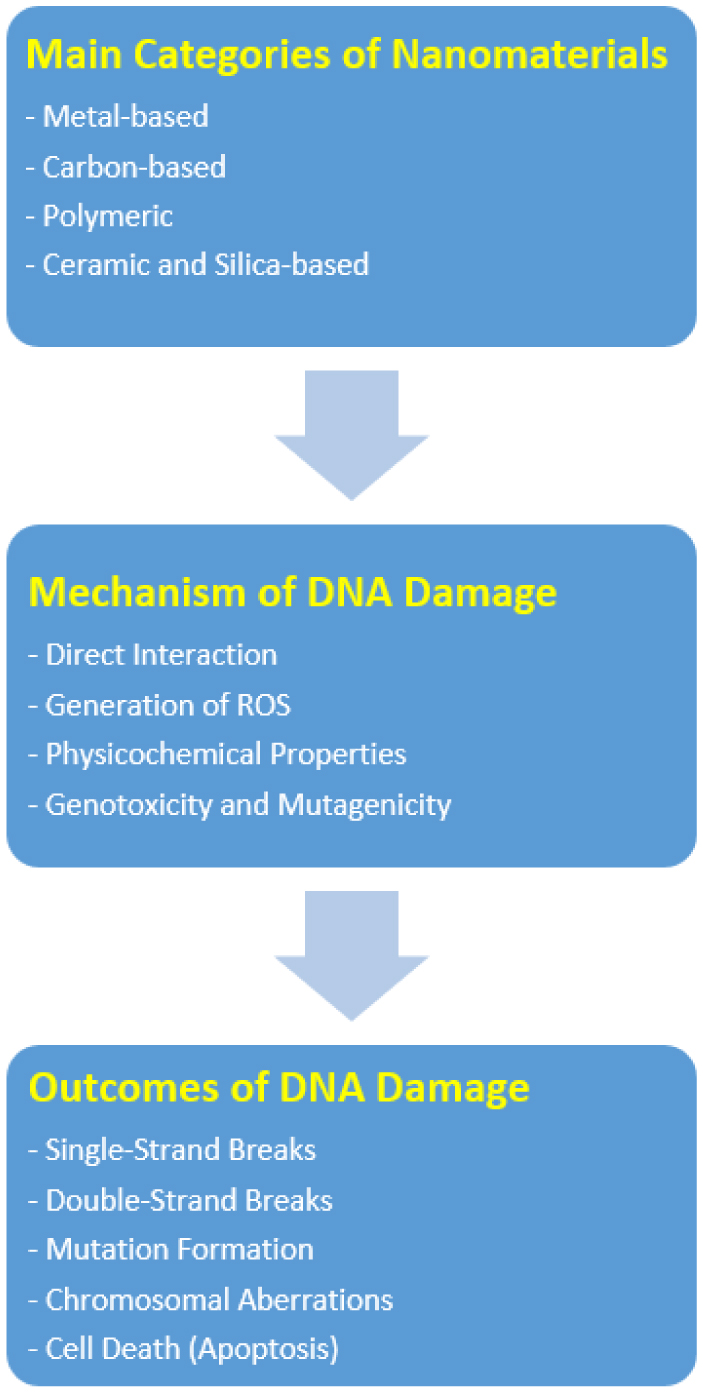
Nanomaterials have garnered significant attention due to their unique properties and wide-ranging applications in medicine and biophysics. However, their interactions with biological systems, particularly DNA, raise critical concerns about genotoxicity and potential long-term health risks. This review delves into the biophysical mechanisms underlying nanomaterial-induced DNA damage, highlighting recent insights, current challenges, and future research directions. We explore how the physicochemical properties of nanomaterials influence their interaction with DNA, the pathways through which they induce damage, and the biophysical methods employed to study these processes.
Citation: James C.L. Chow. Biophysical insights into nanomaterial-induced DNA damage: mechanisms, challenges, and future directions[J]. AIMS Biophysics, 2024, 11(3): 340-369. doi: 10.3934/biophy.2024019
Nanomaterials have garnered significant attention due to their unique properties and wide-ranging applications in medicine and biophysics. However, their interactions with biological systems, particularly DNA, raise critical concerns about genotoxicity and potential long-term health risks. This review delves into the biophysical mechanisms underlying nanomaterial-induced DNA damage, highlighting recent insights, current challenges, and future research directions. We explore how the physicochemical properties of nanomaterials influence their interaction with DNA, the pathways through which they induce damage, and the biophysical methods employed to study these processes.

| [1] | Chow JCL (2017) Application of nanoparticle materials in radiation therapy. Handbook of Ecomaterials . Springer 3661-3681. https://doi.org/10.1007/978-3-319-68255-6_111 |
| [2] | Chow JCL (2020) Recent progress of gold nanomaterials in cancer therapy. Handbook of Nanomaterials and Nanocomposites for Energy and Environmental Applications . Springer 1-30. https://doi.org/10.1007/978-3-030-36268-3_2 |
| [3] |
Dippong T (2024) Innovative nanomaterial properties and applications in chemistry, physics, medicine, or environment. Nanomaterials 14: 145. https://doi.org/10.3390/nano14020145 
|
| [4] |
Yang Z, Chen H, Yang P, et al. (2022) Nano-oxygenated hydrogels for locally and permeably hypoxia relieving to heal chronic wounds. Biomaterials 282: 121401. https://doi.org/10.1016/j.biomaterials.2022.121401
|
| [5] |
Shi L, Song D, Meng C, et al. (2024) Opportunities and challenges of engineered exosomes for diabetic wound healing. Giant 18: 100251. https://doi.org/10.1016/j.giant.2024.100251
|
| [6] |
Fu W, Sun S, Cheng Y, et al. (2024) Opportunities and challenges of nanomaterials in wound healing: Advances, mechanisms, and perspectives. Chem Eng J 495: 153640. https://doi.org/10.1016/j.cej.2024.153640
|
| [7] |
Siddique S, Chow JCL (2020) Application of nanomaterials in biomedical imaging and cancer therapy. Nanomaterials 10: 1700. https://doi.org/10.3390/nano10091700
|
| [8] |
Trucillo P (2024) Biomaterials for drug delivery and human applications. Materials 17: 456. https://doi.org/10.3390/ma17020456
|
| [9] |
Staffurth J (2010) A review of the clinical evidence for intensity-modulated radiotherapy. Clin Oncol 22: 643-657. https://doi.org/10.1016/j.clon.2010.06.013
|
| [10] |
Brito CL, Silva JV, Gonzaga RV, et al. (2024) A review on carbon nanotubes family of nanomaterials and their health field. ACS Omega 9: 8687-8708. https://doi.org/10.1021/acsomega.3c08824
|
| [11] |
Hu J, Dong M (2024) Recent advances in two-dimensional nanomaterials for sustainable wearable electronic devices. J Nanobiotechnol 22: 63. https://doi.org/10.1186/s12951-023-02274-7
|
| [12] |
Rehmanullah MZ, Inayat N, Majeed A (2020) Application of nanoparticles in agriculture as fertilizers and pesticides: challenges and opportunities. New Frontiers in Stress Management for Durable Agriculture : 281-293. https://doi.org/10.1007/978-981-15-1322-0_17
|
| [13] |
Petersen EJ, Nelson BC (2010) Mechanisms and measurements of nanomaterial-induced oxidative damage to DNA. Anal Bioanal Chem 398: 613-650. https://doi.org/10.1007/s00216-010-3881-7
|
| [14] |
Moore JA, Chow JCL (2021) Recent progress and applications of gold nanotechnology in medical biophysics using artificial intelligence and mathematical modeling. Nano Express 2: 022001. https://doi.org/10.1088/2632-959X/abddd3
|
| [15] |
Barua S, Mitragotri S (2014) Challenges associated with penetration of nanoparticles across cell and tissue barriers: a review of current status and future prospects. Nano Today 9: 223-243. https://doi.org/10.1016/j.nantod.2014.04.008
|
| [16] |
Yan L, Gu Z, Zhao Y (2013) Chemical mechanisms of the toxicological properties of nanomaterials: generation of intracellular reactive oxygen species. Chem Asian J 8: 2342-2353. https://doi.org/10.1002/asia.201300542
|
| [17] |
Ruan C, Su K, Zhao D, et al. (2021) Nanomaterials for tumor hypoxia relief to improve the efficacy of ROS-generated cancer therapy. Front Chem 9: 649158. https://doi.org/10.3389/fchem.2021.649158
|
| [18] |
Fu PP, Xia Q, Hwang HM, et al. (2014) Mechanisms of nanotoxicity: generation of reactive oxygen species. J Food Drug Anal 22: 64-75. https://doi.org/10.1016/j.jfda.2014.01.005
|
| [19] | Chow JCL (2016) Photon and electron interactions with gold nanoparticles: a Monte Carlo study on gold nanoparticle-enhanced radiotherapy. Nan Med Imag 8: 45-70. https://doi.org/10.1016/B978-0-323-41736-5.00002-9 |
| [20] |
Chow JCL, Santiago CA (2023) DNA damage of iron-gold nanoparticle heterojunction irradiated by kV photon beams: a Monte Carlo study. Appl Sci 13: 8942. https://doi.org/10.3390/app13158942
|
| [21] |
Santiago CA, Chow JCL (2023) Variations in gold nanoparticle size on DNA damage: a Monte Carlo study based on a multiple-particle model using electron beams. Appl Sci 13: 4916. https://doi.org/10.3390/app13084916
|
| [22] |
Kalyane D, Raval N, Maheshwari R, et al. (2019) Employment of enhanced permeability and retention effect (EPR): nanoparticle-based precision tools for targeting of therapeutic and diagnostic agent in cancer. Mater Sci Eng C 98: 1252-1276. https://doi.org/10.1016/j.msec.2019.01.066
|
| [23] |
Martelli S, Chow JCL (2020) Dose enhancement for the flattening-filter-free and flattening-filter X-ray beams in nanoparticle-enhanced radiotherapy: a Monte Carlo phantom study. Nanomaterials 10: 637. https://doi.org/10.3390/nano10040637
|
| [24] |
Chow JCL (2022) Special issue: application of nanomaterials in biomedical imaging and cancer therapy. Nanomaterials 12: 726. https://doi.org/10.3390/nano12050726
|
| [25] |
Thongkumkoon P, Sangwijit K, Chaiwong C, et al. (2014) Direct nanomaterial-DNA contact effects on DNA and mutation induction. Toxicol Lett 226: 90-97. https://doi.org/10.1016/j.toxlet.2014.01.036
|
| [26] |
Bhabra G, Sood A, Fisher B, et al. (2009) Nanoparticles can cause DNA damage across a cellular barrier. Nat Nanotechnol 4: 876-883. https://doi.org/10.1038/nnano.2009.313
|
| [27] |
Wan R., Mo Y, Feng L, et al. (2012) DNA damage caused by metal nanoparticles: involvement of oxidative stress and activation of ATM. Chem Res Toxicol 25: 1402-1411. https://doi.org/10.1021/tx200513t
|
| [28] |
Zijno A, De Angelis I, De Berardis B, et al. (2015) Different mechanisms are involved in oxidative DNA damage and genotoxicity induction by ZnO and TiO2 nanoparticles in human colon carcinoma cells. Toxicol Vitrp 29: 1503-1512. https://doi.org/10.1016/j.tiv.2015.06.009
|
| [29] |
Hahm JY, Park J, Jang ES, et al. (2022) 8-Oxoguanine: from oxidative damage to epigenetic and epitranscriptional modification. Exp Mol Med 54: 1626-1642. https://doi.org/10.1038/s12276-022-00822-z
|
| [30] |
Letavayová L, Marková E, Hermanská K, et al. (2006) Relative contribution of homologous recombination and non-homologous end-joining to DNA double-strand break repair after oxidative stress in Saccharomyces cerevisiae. DNA Repair 5: 602-610. https://doi.org/10.1016/j.dnarep.2006.01.004
|
| [31] |
Cadet J, Douki T, Gasparutto D, et al. (2003) Oxidative damage to DNA: formation, measurement, and biochemical features. Mutat Res/Fund Mol M 531: 5-23. https://doi.org/10.1016/j.mrfmmm.2003.09.001
|
| [32] |
Encinas-Gimenez M, Martin-Duque P, Martín-Pardillos A (2024) Cellular alterations due to direct and indirect interaction of nanomaterials with nucleic acids. Int J Mol Sci 25: 1983. https://doi.org/10.3390/ijms25041983
|
| [33] |
Li X, Liu W, Sun L, et al. (2015) Effects of physicochemical properties of nanomaterials on their toxicity. J Biomed Mater Res A 103: 2499-2507. https://doi.org/10.1002/jbm.a.35384
|
| [34] |
Li Y, Lian Y, Zhang LT, et al. (2016) Cell and nanoparticle transport in tumour microvasculature: the role of size, shape and surface functionality of nanoparticles. Interface Focus 6: 20150086. https://doi.org/10.1098/rsfs.2015.0086
|
| [35] |
Schaeublin NM, Braydich-Stolle LK, Schrand AM, et al. (2011) Surface charge of gold nanoparticles mediates mechanism of toxicity. Nanoscale 3: 410-420. https://doi.org/10.1039/C0NR00478B
|
| [36] |
Siddique S, Chow JCL (2022) Recent advances in functionalized nanoparticles in cancer theranostics. Nanomaterials 12: 2826. https://doi.org/10.3390/nano12162826
|
| [37] |
Singh N, Manshian B, Jenkins GJ, et al. (2009) NanoGenotoxicology: the DNA damaging potential of engineered nanomaterials. Biomaterials 30: 3891-3914. https://doi.org/10.1016/j.biomaterials.2009.04.009
|
| [38] |
Landsiedel R, Honarvar N, Seiffert SB, et al. (2022) Genotoxicity testing of nanomaterials. WIRES: Nanomed Nanobiotechnol 14: e1833. https://doi.org/10.1002/wnan.1833
|
| [39] |
Møller P, Roursgaard M (2024) Gastrointestinal tract exposure to particles and DNA damage in animals: a review of studies before, during, and after the peak of nanotoxicology. Mut Res/Rev Mutat Res 793: 108491. https://doi.org/10.1016/j.mrrev.2024.108491
|
| [40] |
Chow JCL (2021) Synthesis and applications of functionalized nanoparticles in biomedicine and radiotherapy. Additive Manufacturing with Functionalized Nanomaterials . Elsevier 193-214. https://doi.org/10.1016/B978-0-12-823152-4.00001-6
|
| [41] |
Chompoosor A, Saha K, Ghosh PS, et al. (2010) The role of surface functionality on acute cytotoxicity, ROS generation and DNA damage by cationic gold nanoparticles. Small (Weinheim an der Bergstrasse, Germany) 6: 2246. https://doi.org/10.1002/smll.201000463
|
| [42] |
Carlson C, Hussain SM, Schrand AM, et al. (2008) Unique cellular interaction of silver nanoparticles: size-dependent generation of reactive oxygen species. J Phys Chem B 112: 13608-13619. https://doi.org/10.1021/jp712087m
|
| [43] |
Song MF, Li YS, Kasai H, et al. (2012) Metal nanoparticle-induced micronuclei and oxidative DNA damage in mice. J Clin Biochem Nutr 50: 211-216. https://doi.org/10.3164/jcbn.11-70
|
| [44] |
Sotiropoulos M, Henthorn NT, Warmenhoven JW, et al. (2017) Modelling direct DNA damage for gold nanoparticle enhanced proton therapy. Nanoscale 9: 18413-18422. https://doi.org/10.1039/C7NR07310K
|
| [45] |
Madannejad R, Shoaie N, Jahanpeyma F, et al. (2019) Toxicity of carbon-based nanomaterials: reviewing recent reports in medical and biological systems. Chem-Biol Interact 307: 206-222. https://doi.org/10.1016/j.cbi.2019.04.036
|
| [46] |
Heredia DA, Durantini AM, Durantini JE, et al. (2022) Fullerene C60 derivatives as antimicrobial photodynamic agents. J Photochem Photobiol Photochem Rev 51: 100471. https://doi.org/10.1016/j.jphotochemrev.2021.100471
|
| [47] |
Migliore L, Saracino D, Bonelli A, et al. (2010) Carbon nanotubes induce oxidative DNA damage in RAW 264.7 cells. Environ Mol Mutagen 51: 294-303. https://doi.org/10.1002/em.20545
|
| [48] |
Oh WK, Kwon OS, Jang J (2013) Conducting polymer nanomaterials for biomedical applications: cellular interfacing and biosensing. Polym Rev 53: 407-442. https://doi.org/10.1080/15583724.2013.805771
|
| [49] | Kulkarni AA, Rao PS (2013) Synthesis of polymeric nanomaterials for biomedical applications. Nanomaterials in Tissue Engineering . Woodhead Publishing 27-63. https://doi.org/10.1533/9780857097231.1.27 |
| [50] |
Li J, Pu K (2020) Semiconducting polymer nanomaterials as near-infrared photoactivatable protherapeutics for cancer. Acc Chem Res 53: 752-762. https://doi.org/10.1021/acs.accounts.9b00569
|
| [51] | Balasubramanian SB, Gurumurthy B, Balasubramanian A (2017) Biomedical applications of ceramic nanomaterials: a review. Int J Pharm Sci Res 8: 4950-4959. https://doi.org/10.13040/IJPSR.0975-8232.8(12).4950-59 |
| [52] |
Jafari S, Mahyad B, Hashemzadeh H, et al. (2020) Biomedical applications of TiO2 nanostructures: recent advances. Int J Nanomed 15: 3447-3470. https://doi.org/10.2147/IJN.S249441
|
| [53] |
Huang Y, Li P, Zhao R, et al. (2022) Silica nanoparticles: biomedical applications and toxicity. Biomed Pharmacother 151: 113053. https://doi.org/10.1016/j.biopha.2022.113053
|
| [54] |
Chen L, Liu J, Zhang Y, et al. (2018) The toxicity of silica nanoparticles to the immune system. Nanomedicine 13: 1939-1962. https://doi.org/10.2217/nnm-2018-0076
|
| [55] |
Dolai J, Mandal K, Jana NR (2021) Nanoparticle size effects in biomedical applications. ACS Appl Nano Mater 4: 6471-6496. https://doi.org/10.1021/acsanm.1c00987
|
| [56] |
Albanese A, Tang PS, Chan WC (2012) The effect of nanoparticle size, shape, and surface chemistry on biological systems. Annu Rev Biomed Eng 14: 1-6. https://doi.org/10.1146/annurev-bioeng-071811-150124
|
| [57] |
Yang H, Liu C, Yang D, et al. (2009) Comparative study of cytotoxicity, oxidative stress, and genotoxicity induced by four typical nanomaterials: the role of particle size, shape, and composition. J Appl Toxicol 29: 69-78. https://doi.org/10.1002/jat.1385
|
| [58] |
Khaing Oo MK, Yang Y, Hu Y, et al. (2012) Gold nanoparticle-enhanced and size-dependent generation of reactive oxygen species from protoporphyrin IX. ACS Nano 6: 1939-1947. https://doi.org/10.1021/nn300327c
|
| [59] |
Kang Z, Yan X, Zhao L, et al. (2015) Gold nanoparticle/ZnO nanorod hybrids for enhanced reactive oxygen species generation and photodynamic therapy. Nano Res 8: 2004-2014. https://doi.org/10.1007/s12274-015-0712-3
|
| [60] |
Subbiah R, Veerapandian M, Yun KS (2010) Nanoparticles: Functionalization and multifunctional applications in biomedical sciences. Curr Med Chem 17: 4559-4577. https://doi.org/10.2174/092986710794183024
|
| [61] |
Fröhlich E (2012) The role of surface charge in cellular uptake and cytotoxicity of medical nanoparticles. Int J Nanomed 7: 5577-5591. https://doi.org/10.2147/IJN.S36111
|
| [62] |
Siddique S, Chow JCL (2020) Gold nanoparticles for drug delivery and cancer therapy. Appl Sci 10: 3824. https://doi.org/10.3390/app10113824
|
| [63] |
Suk JS, Xu Q, Kim N, et al. (2016) PEGylation as a strategy for improving nanoparticle-based drug and gene delivery. Adv Drug Deliv Rev 99: 28-51. https://doi.org/10.1016/j.addr.2015.09.012
|
| [64] |
Shi M, Kwon HS, Peng Z, et al. (2012) Effects of surface chemistry on the generation of reactive oxygen species by copper nanoparticles. ACS Nano 6: 2157-2164. https://doi.org/10.1021/nn300445d
|
| [65] |
Čapek J, Roušar T (2021) Detection of oxidative stress induced by nanomaterials in cells—the roles of reactive oxygen species and glutathione. Molecules 26: 4710. https://doi.org/10.3390/molecules26164710
|
| [66] |
Magdolenova Z, Bilaničová D, Pojana G, et al. (2012) Impact of agglomeration and different dispersions of titanium dioxide nanoparticles on the human related in vitro cytotoxicity and genotoxicity. J Environ Monitor 14: 455-464. https://doi.org/10.1039/C2EM10746E
|
| [67] |
Behzadi S, Serpooshan V, Tao W, et al. (2017) Cellular uptake of nanoparticles: journey inside the cell. Chem Soc Rev 46: 4218-4244. https://doi.org/10.1039/C6CS00636A
|
| [68] |
Soto K, Garza KM, Murr LE (2007) Cytotoxic effects of aggregated nanomaterials. Acta Biomater 3: 351-358. https://doi.org/10.1016/j.actbio.2006.11.004
|
| [69] |
Liu Y, Zhu S, Gu Z, et al. (2022) Toxicity of manufactured nanomaterials. Particuology 69: 31-48. https://doi.org/10.1016/j.partic.2021.11.007
|
| [70] |
Walkey CD, Chan WC (2012) Understanding and controlling the interaction of nanomaterials with proteins in a physiological environment. Chem Soc Rev 41: 2780-2799. https://doi.org/10.1039/C1CS15233E
|
| [71] | Lee YK, Choi EJ, Webster TJ, et al. (2015) Effect of the protein corona on nanoparticles for modulating cytotoxicity and immunotoxicity. Int J Nanomed 10: 97-113. https://doi.org/10.2147/IJN.S72998 |
| [72] |
Bushell M, Beauchemin S, Kunc F, et al. (2020) Characterization of commercial metal oxide nanomaterials: Crystalline phase, particle size, and specific surface area. Nanomaterials 10: 1812. https://doi.org/10.3390/nano10091812
|
| [73] |
Mahaye N, Thwala M, Cowan DA, et al. (2017) Genotoxicity of metal-based engineered nanoparticles in aquatic organisms: a review. Mut Res/Rev Mut Res 773: 134-160. https://doi.org/10.1016/j.mrrev.2017.05.004
|
| [74] |
Zijno A, De Angelis I, De Berardis B, et al. (2015) Different mechanisms are involved in oxidative DNA damage and genotoxicity induction by ZnO and TiO2 nanoparticles in human colon carcinoma cells. Toxicol Vitro 29: 1503-1512. https://doi.org/10.1016/j.tiv.2015.06.009
|
| [75] |
Racovita AD (2022) Titanium dioxide: structure, impact, and toxicity. Int J Environ Res Public Health 19: 5681. https://doi.org/10.3390/ijerph19095681
|
| [76] |
Sukhanova A, Bozrova S, Sokolov P, et al. (2018) Dependence of nanoparticle toxicity on their physical and chemical properties. Nanoscale Res Lett 13: 44. https://doi.org/10.1186/s11671-018-2457-x
|
| [77] |
Thu HE, Haider MA, Khan S, et al. (2023) Nanotoxicity induced by nanomaterials: a review of factors affecting nanotoxicity and possible adaptations. OpenNano 14: 100190. https://doi.org/10.1016/j.onano.2023.100190
|
| [78] |
Sirajuddin M, Ali S, Badshah A (2013) Drug–DNA interactions and their study by UV–Visible, fluorescence spectroscopies, and cyclic voltammetry. J Photoch Photobio B 124: 1-9. https://doi.org/10.1016/j.jphotobiol.2013.03.013
|
| [79] |
Wamsley M, Zou S, Zhang D (2023) Advancing evidence-based data interpretation in UV–Vis and fluorescence analysis for nanomaterials: an analytical chemistry perspective. Anal Chem 95: 17426-17437. https://doi.org/10.1021/acs.analchem.3c03490
|
| [80] |
Suh JS, Kim TJ (2023) A novel DNA double-strand breaks biosensor based on fluorescence resonance energy transfer. Biomater Res 27: 15. https://doi.org/10.1186/s40824-023-00354-1
|
| [81] |
Kolyvanova MA, Klimovich MA, Belousov AV, et al. (2022) A principal approach to the detection of radiation-induced DNA damage by circular dichroism spectroscopy and its dosimetric application. Photonics 9: 787. https://doi.org/10.3390/photonics9110787
|
| [82] |
Xu X, Nakano T, Tsuda M, et al. (2020) Direct observation of damage clustering in irradiated DNA with atomic force microscopy. Nucleic Acids Res 48: e18. https://doi.org/10.1093/nar/gkz1159
|
| [83] |
Rübe CE, Lorat Y, Schuler N, et al. (2011) DNA repair in the context of chromatin: new molecular insights by the nanoscale detection of DNA repair complexes using transmission electron microscopy. DNA Repair 10: 427-437. https://doi.org/10.1016/j.dnarep.2011.01.012
|
| [84] |
Scalisi S, Privitera AP, Pelicci PG, et al. (2024) Origin and evolution of oncogene-related DNA damage: a confocal imaging study. Biophys J 123: 290a-291a. https://doi.org/10.1016/j.bpj.2023.11.1811
|
| [85] |
Darwanto A, Farrel A, Rogstad DK, et al. (2009) Characterization of DNA glycosylase activity by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry. Anal Biochem 394: 13-23. https://doi.org/10.1016/j.ab.2009.07.015
|
| [86] |
Chaudhary AK, Nokubo M, Oglesby TD, et al. (1995) Characterization of endogenous DNA adducts by liquid chromatography/electrospray ionization tandem mass spectrometry. J Mass Spectrom 30: 1157-1166. https://doi.org/10.1002/jms.1190300813
|
| [87] |
Kaneko S, Takamatsu K (2024) Angle modulated two-dimensional single cell pulsed-field gel electrophoresis for detecting early symptoms of DNA fragmentation in human sperm nuclei. Sci Rep 14: 840. https://doi.org/10.1038/s41598-024-51509-6
|
| [88] |
Plitta-Michalak BP, Ramos A, Stępień D, et al. (2024) Pespective: the comet assay as a method for assessing DNA damage in cryopreserved samples. CryoLetters 45: 1-5. https://doi.org/10.54680/fr24110110112
|
| [89] |
Chatha AMM, Naz S, Iqbal SS, et al. (2024) Detection of DNA damage in fish using comet assay. Curr Trends in OMICS 4: 01-16. https://doi.org/10.32350/cto.41.01
|
| [90] |
Li H, Xu Y, Shi W, et al. (2017) Assessment of alterations in X-ray irradiation-induced DNA damage of glioma cells by using proton nuclear magnetic resonance spectroscopy. Int J Biochem Cell Biol 84: 109-118. https://doi.org/10.1016/j.biocel.2017.01.010
|
| [91] |
Campagne S, Gervais V, Milon A (2011) Nuclear magnetic resonance analysis of protein–DNA interactions. J R Soc Interface 8: 1065-1078. https://doi.org/10.1098/rsif.2010.0543
|
| [92] |
Abolfath RM, Carlson DJ, Chen ZJ, et al. (2013) A molecular dynamics simulation of DNA damage induction by ionizing radiation. Phys Med Biol 58: 7143. https://doi.org/10.1088/0031-9155/58/20/7143
|
| [93] |
Yang S, Zhao T, Zou L, et al. (2019) ReaxFF-based molecular dynamics simulation of DNA molecules destruction in cancer cells by plasma ROS. Phys Plasmas 26: 083504. https://doi.org/10.1063/1.5097243
|
| [94] |
Sheeraz Z, Chow JCL (2021) Evaluation of dose enhancement with gold nanoparticles in kilovoltage radiotherapy using the new EGS geometry library in Monte Carlo simulation. AIMS Biophys 8: 337-345. https://doi.org/10.3934/biophy.2021027
|
| [95] |
Leung MK, Chow JC, Chithrani BD, et al. (2011) Irradiation of gold nanoparticles by x-rays: Monte Carlo simulation of dose enhancements and the spatial properties of the secondary electrons production. Med Phys 38: 624-631. https://doi.org/10.1118/1.3539623
|
| [96] | Chow JCL (2018) Monte Carlo nanodosimetry in gold nanoparticle-enhanced radiotherapy. Recent Advancements and Applications in Dosimetry . New York: Nova Science Publishers. |
| [97] |
Jabeen M, Chow JCL (2021) Gold nanoparticle DNA damage by photon beam in a magnetic field: a Monte Carlo study. Nanomaterials 11: 1751. https://doi.org/10.3390/nano11071751
|
| [98] |
Chun H, Chow JCL (2016) Gold nanoparticle DNA damage in radiotherapy: a Monte Carlo study. AIMS Bioeng 3: 352-361. https://doi.org/10.3934/bioeng.2016.3.352
|
| [99] |
Horvath T, Papp A, Igaz N, et al. (2018) Pulmonary impact of titanium dioxide nanorods: examination of nanorod-exposed rat lungs and human alveolar cells. Int J Nanomed 13: 7061-7077. https://doi.org/10.2147/IJN.S179159
|
| [100] |
AshaRani PV, Low Kah Mun G, Hande MP, et al. (2009) Cytotoxicity and genotoxicity of silver nanoparticles in human cells. ACS Nano 3: 279-290. https://doi.org/10.1021/nn800596w
|
| [101] |
Karlsson HL, Cronholm P, Gustafsson J, et al. (2008) Copper oxide nanoparticles are highly toxic: a comparison between metal oxide nanoparticles and carbon nanotubes. Chem Res Toxicol 21: 1726-1732. https://doi.org/10.1021/tx800064j
|
| [102] |
Singh N, Manshian B, Jenkins GJ, et al. (2009) NanoGenotoxicology: the DNA damaging potential of engineered nanomaterials. Biomaterials 30: 3891-3914. https://doi.org/10.1016/j.biomaterials.2009.04.009
|
| [103] |
Magdolenova Z, Collins A, Kumar A, et al. (2014) Mechanisms of genotoxicity: a review of in vitro and in vivo studies with engineered nanoparticles. Nanotoxicology 8: 233-278. https://doi.org/10.3109/17435390.2013.773464
|
| [104] |
Gonzalez L, Lison D, Kirsch-Volders M (2008) Genotoxicity of engineered nanomaterials: a critical review. Nanotoxicology 2: 252-273. https://doi.org/10.1080/17435390802464986
|
| [105] |
Ahamed M, Karns M, Goodson M, et al. (2008) DNA damage response to different surface chemistry of silver nanoparticles in mammalian cells. Toxicol Appl Pharm 233: 404-410. https://doi.org/10.1016/j.taap.2008.09.015
|
| [106] |
Shukla RK, Sharma V, Pandey AK, et al. (2011) ROS-mediated genotoxicity induced by titanium dioxide nanoparticles in human epidermal cells. Toxicol in Vitro 25: 231-241. https://doi.org/10.1016/j.tiv.2010.11.008
|
| [107] |
Sharma V, Singh P, Pandey AK, et al. (2012) Induction of oxidative stress and DNA damage by zinc oxide nanoparticles in human liver cells (HepG2). J Biomed Nanotechnol 8: 63-65. https://doi.org/10.1016/j.mrgentox.2011.12.009
|
| [108] |
Oberdörster G, Oberdörster E, Oberdörster J (2005) Nanotoxicology: an emerging discipline evolving from studies of ultrafine particles. Environ Health Persp 113: 823-839. https://doi.org/10.1289/ehp.7339
|
| [109] |
Park EJ, Yi J, Kim Y, et al. (2010) Silver nanoparticles induce cytotoxicity by a Trojan-horse type mechanism. Toxicol Vitro 24: 872-878. https://doi.org/10.1016/j.tiv.2009.12.001
|
| [110] |
Chen Z, Meng H, Xing G, et al. (2006) Acute toxicological effects of copper nanoparticles in vivo. Toxicol Lett 163: 109-120. https://doi.org/10.1016/j.toxlet.2005.10.003
|
| [111] |
Trouiller B, Reliene R, Westbrook A, et al. (2009) Titanium dioxide nanoparticles induce DNA damage and genetic instability in vivo in mice. Cancer Res 69: 8784-8789. https://doi.org/10.1158/0008-5472.CAN-09-2496
|
| [112] |
Folkmann JK, Risom L, Jacobsen NR, et al. (2009) Oxidatively damaged DNA in rats exposed by oral gavage to C60 fullerenes and single-walled carbon nanotubes. Environ Health Persp 117: 703-708. https://doi.org/10.1289/ehp.11922
|
| [113] |
Bahamonde J, Brenseke B, Prater MR, et al. (2018) Gold nanoparticles toxicity in mice and rats: species differences. Toxicol Pathol 46: 431-443. https://doi.org/10.1177/0192623318770608
|
| [114] | Lam CW, James JT, McCluskey R, et al. (2004) Pulmonary toxicity of single-wall carbon nanotubes in mice 7 and 90 days after intratracheal instillation. Toxicol Sci 77: 26-134. https://doi.org/10.1093/toxsci/kfg243 |
| [115] |
Pan Y, Neuss S, Leifert A, et al. (2007) Size-dependent cytotoxicity of gold nanoparticles. Small 3: 1941-1949. https://doi.org/10.1002/smll.200700378
|
| [116] |
Jiang W, Kim BY, Rutka JT, et al. (2008) Nanoparticle-mediated cellular response is size-dependent. Nat Nanotechnol 3: 145-150. https://doi.org/10.1038/nnano.2008.30
|
| [117] | Zhang XD, Wu D, Shen X, et al. (2012) Size-dependent in vivo toxicity of PEG-coated gold nanoparticles. Int J Nanomed 6: 2071-2081. https://doi.org/10.2147/IJN.S21657 |
| [118] |
Derfus AM, Chan WC, Bhatia SN (2004) Probing the cytotoxicity of semiconductor quantum dots. Nano Lett 4: 11-18. https://doi.org/10.1021/nl0347334
|
| [119] |
Ahamed M, Siddiqui MK, Akhtar MJ, et al. (2010) Genotoxic potential of copper oxide nanoparticles in human lung epithelial cells. Biochem Bioph Res Co 396: 578-583. https://doi.org/10.1016/j.bbrc.2010.04.156
|
| [120] |
Kang S, Pinault M, Pfefferle LD, et al. (2008) Single-walled carbon nanotubes exhibit strong antimicrobial activity. Langmuir 24: 6409-6413. https://doi.org/10.1021/la701067r
|
| [121] |
Limbach LK, Wick P, Manser P, et al. (2007) Exposure of engineered nanoparticles to human lung epithelial cells: Influence of chemical composition and catalytic activity on oxidative stress. Environ Sci Technol 41: 4158-4163. https://doi.org/10.1021/es062629t
|
| [122] |
Collins AR (2004) The comet assay for DNA damage and repair: principles, applications, and limitations. Mol Biotechnol 26: 249-261. https://doi.org/10.1385/MB:26:3:249
|
| [123] |
Fenech M (2000) The in vitro micronucleus technique. Mutat Res/Fund Mol M 455: 81-95. https://doi.org/10.1016/S0027-5107(00)00065-8
|
| [124] |
Rogakou EP, Pilch DR, Orr AH, et al. (1998) DNA double-stranded breaks induce histone H2AX phosphorylation on serine 139. J Biol Chem 273: 5858-5868. https://doi.org/10.1074/jbc.273.10.5858
|
| [125] |
Olive PL, Banáth JP (2006) The comet assay: a method to measure DNA damage in individual cells. Nat Protoc 1: 23-29. https://doi.org/10.1038/nprot.2006.5
|
| [126] |
Kirsch-Volders M, Sofuni T, Aardema M, et al. (2011) Report from the in vitro micronucleus assay working group. Mut Res/Genet Toxicol Environ Mutagen 540: 153-163. https://doi.org/10.1016/j.mrgentox.2003.07.005
|
| [127] |
Mah LJ, El-Osta A, Karagiannis TC (2010) γH2AX: a sensitive molecular marker of DNA damage and repair. Leukemia 24: 679-686. https://doi.org/10.1038/leu.2010.6
|
| [128] |
AshaRani PV, Low Kah Mun G, Hande MP, et al. (2009) Cytotoxicity and genotoxicity of silver nanoparticles. ACS Nano 3: 279-290. https://doi.org/10.1021/nn800596w
|
| [129] |
Gurr JR, Wang AS, Chen CH, et al. (2005) Ultrafine titanium dioxide particles in the absence of photoactivation can induce oxidative damage to human bronchial epithelial cells. Toxicology 213: 66-73. https://doi.org/10.1016/j.tox.2005.05.007
|
| [130] |
Migliore L, Saracino S, Bonfiglioli R, et al. (2010) Carbon nanotubes induce oxidative DNA damage in RAW264.7 cells. Environ Mol Mutagen 51: 294-303. https://doi.org/10.1002/em.20545
|
| [131] | Tsuchiya T, Oguri I, Yamakoshi YN, et al. (1996) Novel harmful effects of [60] fullerene on mouse embryos in vitro and in vivo. EBS Lett 393: 139-145. https://doi.org/10.1016/0014-5793(96)00812-5 |
| [132] |
Gupta SK, Sundarraj K, Devashya N, et al. (2013) ZnO nanoparticles induce apoptosis in human dermal fibroblasts via p53-p21 mediated ROS generation and mitochondrial oxidative stress. Biotechnol Bioeng 110: 3113-3122. https://doi.org/10.1016/j.tiv.2011.08.011
|
| [133] |
Chow JCL (2018) Recent progress in Monte Carlo simulation on gold nanoparticle radiosensitization. AIMS Biophys 5: 231-244. https://doi.org/10.3934/biophy.2018.4.231
|
| [134] |
Chithrani DB, Jelveh S, Jalali F, et al. (2010) Gold nanoparticles as radiation sensitizers in cancer therapy. Radiat Res 173: 719-728. https://doi.org/10.1667/RR1984.1
|
| [135] |
Zheng XJ, Chow JCL (2017) Radiation dose enhancement in skin therapy with nanoparticle addition: a Monte Carlo study on kilovoltage photon and megavoltage electron beams. World J Radiol 9: 63-71. https://doi.org/10.4329/wjr.v9.i2.63
|
| [136] |
Chow JCL (2020) Depth dose enhancement on flattening-filter-free photon beam: a Monte Carlo study in nanoparticle-enhanced radiotherapy. Appl Sci 10: 7052. https://doi.org/10.3390/app10207052
|
| [137] |
Cho SH, Jones BL, Krishnan S (2005) The dosimetric feasibility of gold nanoparticle-aided radiation therapy (GNRT) via brachytherapy using low-energy gamma-/X-ray sources. Phys Med Biol 50: N163-N173. https://doi.org/10.1088/0031-9155/54/16/004
|
| [138] |
Cho S, Jeong JH, Kim CH, et al. (2010) Monte Carlo simulation study on dose enhancement by gold nanoparticles in brachytherapy. J Korean Phys Soc 56: 1754-1758. https://doi.org/10.3938/jkps.56.1754
|



Figures(4) / Tables(1)

James C.L. Chow. Biophysical insights into nanomaterial-induced DNA damage: mechanisms, challenges, and future directions[J]. AIMS Biophysics, 2024, 11(3): 340-369. doi: 10.3934/biophy.2024019







 DownLoad:
DownLoad: