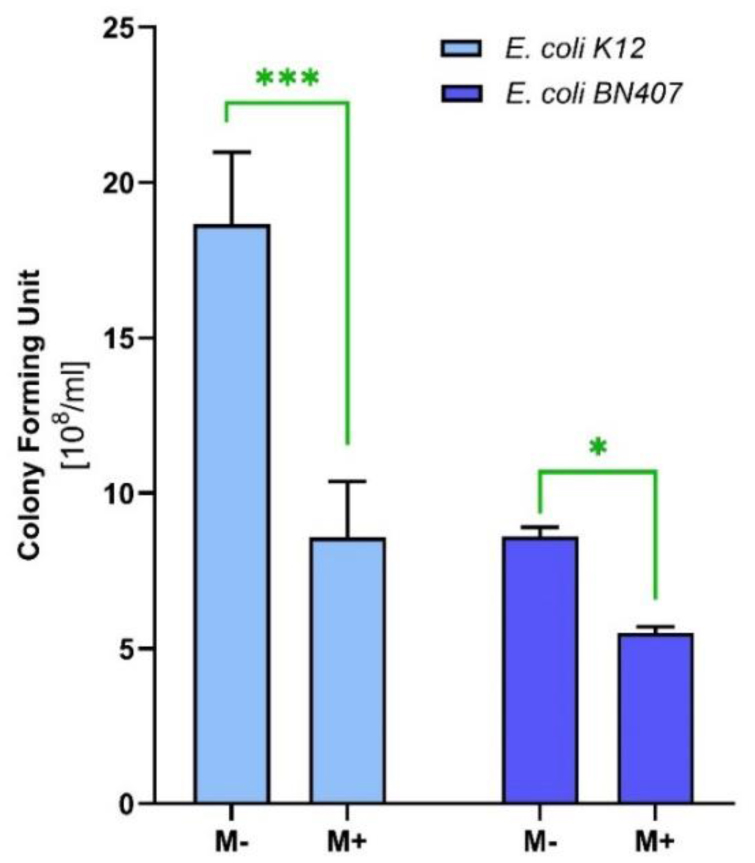
In this investigation, we assessed the antibacterial properties of menthol using Escherichia coli strains, including a tetracycline-resistant (BN407) and a non-resistant reference (K12) strains. The disc-diffusion assay indicated remarkable antibacterial activity of menthol, warranting further exploration. The minimum inhibitory concentration (MIC) for menthol was determined to be 500 µg/ml for both strains. Despite identical MIC values, menthol exhibited different effects on the colony-forming units (CFUs) of the strains, reducing CFUs by 55% in E. coli K12 and 40% in E. coli BN407.
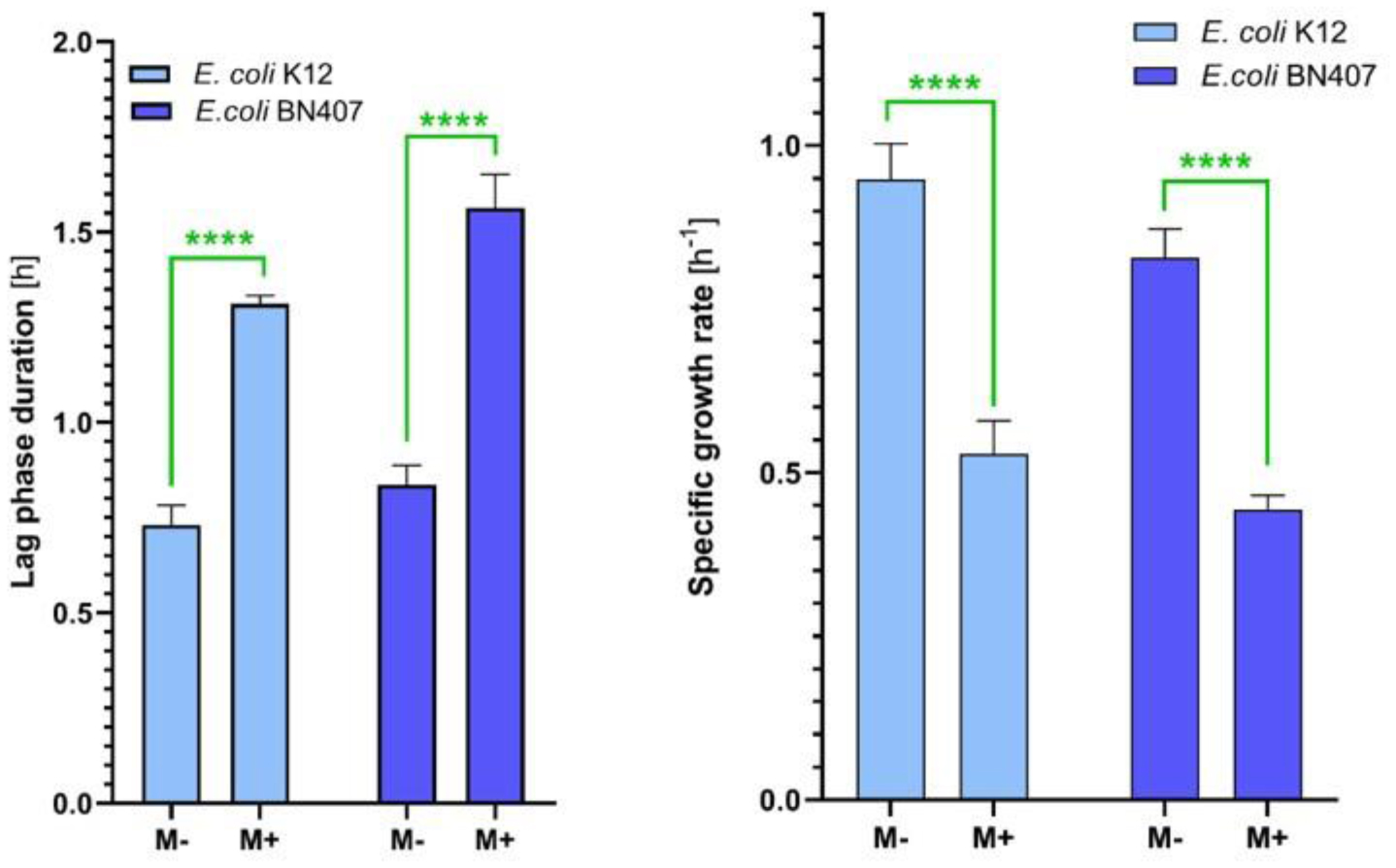
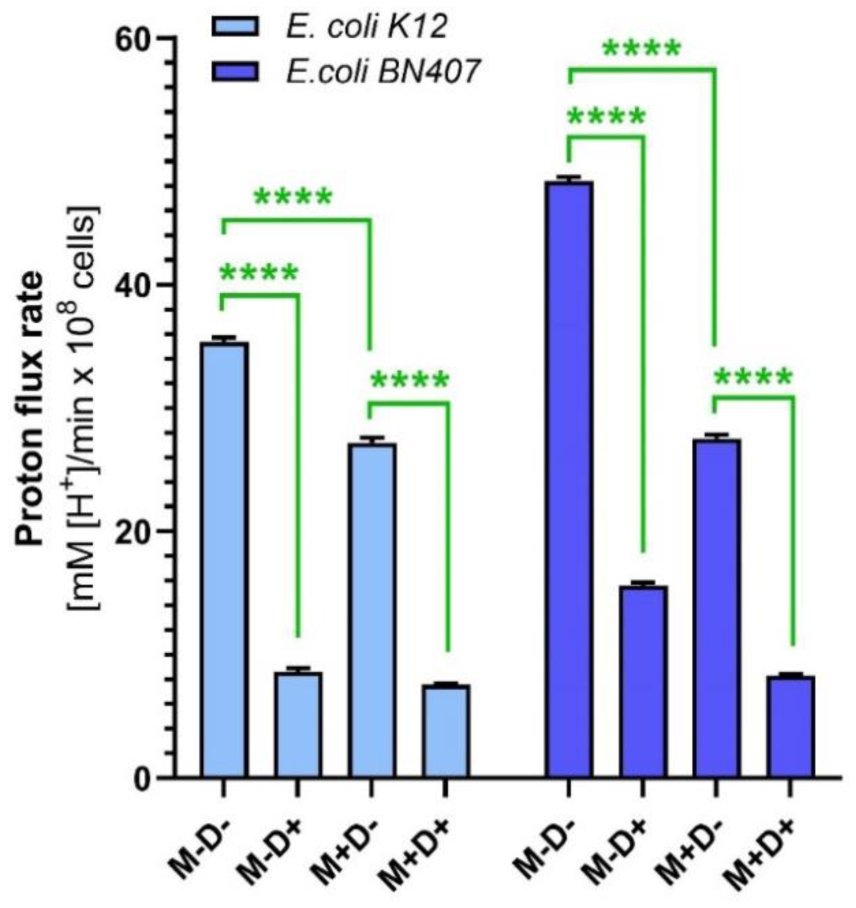
Growth kinetics studies revealed that menthol extended the Lag phase by 50% and decreased the specific growth rate and mean generation time by nearly 50% for both strains. These findings illustrate menthol's significant impact on bacterial replication and adaptation processes. Additionally, menthol disrupted membrane-associated properties, as evidenced by reduced H+-flux through bacterial membranes, affecting both N,N-Dicyclohexylcarbodiimide (DCCD)-sensitive and non-sensitive proton flux rates. This indicates that menthol compromises the proton motive force critical for ATP synthesis and nutrient transport.
In summary, menthol demonstrates potent antibacterial activity, influencing bacterial growth and survival. This activity is supposed to be due to the influence on membrane functionality. These effects are consistent across both tetracycline-resistant and non-resistant E. coli strains.
Citation: Silvard Tadevosyan, Naira Sahakyan. Influence of menthol on membrane-associated properties of tetracycline-resistant Escherichia coli[J]. AIMS Biophysics, 2024, 11(3): 329-339. doi: 10.3934/biophy.2024018
In this investigation, we assessed the antibacterial properties of menthol using Escherichia coli strains, including a tetracycline-resistant (BN407) and a non-resistant reference (K12) strains. The disc-diffusion assay indicated remarkable antibacterial activity of menthol, warranting further exploration. The minimum inhibitory concentration (MIC) for menthol was determined to be 500 µg/ml for both strains. Despite identical MIC values, menthol exhibited different effects on the colony-forming units (CFUs) of the strains, reducing CFUs by 55% in E. coli K12 and 40% in E. coli BN407.
Growth kinetics studies revealed that menthol extended the Lag phase by 50% and decreased the specific growth rate and mean generation time by nearly 50% for both strains. These findings illustrate menthol's significant impact on bacterial replication and adaptation processes. Additionally, menthol disrupted membrane-associated properties, as evidenced by reduced H+-flux through bacterial membranes, affecting both N,N-Dicyclohexylcarbodiimide (DCCD)-sensitive and non-sensitive proton flux rates. This indicates that menthol compromises the proton motive force critical for ATP synthesis and nutrient transport.
In summary, menthol demonstrates potent antibacterial activity, influencing bacterial growth and survival. This activity is supposed to be due to the influence on membrane functionality. These effects are consistent across both tetracycline-resistant and non-resistant E. coli strains.
essential oil
compound annual growth rate
Specific growth rate
wild type
standard deviation
minimal inhibitory concentration
gas chromatography mass spectrometry

| [1] |
Seukep AJ, Mbuntcha HG, Zeuko'o EM, et al. (2023) Established antibacterial drugs from plants. Adv Bot Res 106: 81-149. https://doi.org/10.1016/bs.abr.2022.08.005 
|
| [2] |
Vaou N, Stavropoulou E, Voidarou C, et al. (2021) Advances in medicinal plant antimicrobial activity: a review study on challenges and future perspectives. Microorganisms 9: 2041. https://doi.org/10.3390/microorganisms9102041
|
| [3] |
Ginovyan M, Babayan A, Shirvanyan A, et al. (2023) The action mechanisms, anti-cancer and antibiotic-modulation potential of Vaccinium myrtillus L. extract. Discov Med 35: 590-611. https://doi.org/10.24976/Discov.Med.202335177.59
|
| [4] |
Vaou N, Stavropoulou E, Voidarou CC, et al. (2022) Interactions between medical plant-derived bioactive compounds: focus on antimicrobial combination effects. Antibiotics (Basel) 11: 1014. https://doi.org/10.3390/antibiotics11081014
|
| [5] |
Imlay JA (2019) Where in the world do bacteria experience oxidative stress?. Environ Microbiol 21: 521-530. https://doi.org/10.1111/1462-2920.14445
|
| [6] |
Seukep AJ, Kuete V, Nahar L, et al. (2020) Plant-derived secondary metabolites as the main source of efflux pump inhibitors and methods for identification. J Pharm Anal 10: 277-290. https://doi.org/10.1016/j.jpha.2019.11.002
|
| [7] | Sahakyan N, Petrosyan M, Trchounian A (2019) Some Lamiaceae family plant essential oil chemical composition and their potential as antimicrobial agents against antibiotic-resistant bacteria. Proceedings of the YSU B: Chemical and Biological Sciences 53: 23-28. https://doi.org/10.46991/PYSU:B/2019.53.1.023 |
| [8] |
Sahakyan N (2023) Lamiaceae family plants: one of the potentially richest sources of antimicrobials. Pharm Chem J 57: 507-5014. https://doi.org/10.1007/s11094-023-02921-1
|
| [9] | Salehi B, Stojanović-Radić Z, Matejić J, et al. (2018) Plants of genus Mentha: from farm to food factory. Plants (Basel) 7: 70. https://doi.org/10.3390/plants7030070 |
| [10] |
Landau E, Shapira R (2012) Effects of subinhibitory concentrations of menthol on adaptation, morphological, and gene expression changes in enterohemorrhagic Escherichia coli. Appl Environ Microbiol 78: 5361-5367. https://doi.org/10.1128/AEM.00894-12
|
| [11] |
Nazzaro F, Fratianni F, De Martino L, et al. (2013) Effect of essential oils on pathogenic bacteria. Pharmaceuticals (Basel) 6: 1451-1474. https://doi.org/10.3390/ph6121451
|
| [12] |
Fernández-Billón M, Llambías-Cabot AE, Jordana-Lluch E, et al. (2023) Mechanisms of antibiotic resistance in Pseudomonas aeruginosa biofilms. Biofilm 5: 100129. https://doi.org/10.1016/j.bioflm.2023.100129
|
| [13] |
Husain FM, Ahmad I, Khan MS, et al. (2015) Sub-MICs of Mentha piperita essential oil and menthol inhibits AHL mediated quorum sensing and biofilm of gram-negative bacteria. Front Microbiol 6: 420. https://doi.org/10.3389/fmicb.2015.00420
|
| [14] |
Ferreira de Melo AN, Targino de Souza Pedrosa G, Tayse da Cruz Almeida E, et al. (2020) Successive exposure to Mentha piperita L. essential oil affects the culturability and induces membrane repair in a persister epidemic Salmonella Typhimurium PT4. Microb Pathog 149: 104264. https://doi.org/10.1016/j.micpath.2020.104264
|
| [15] |
Trombetta D, Castelli F, Sarpietro MG, et al. (2005) Mechanisms of antibacterial action of three monoterpenes. Antimicrob Agents Ch 49: 2474-2478. https://doi.org/10.1128/aac.49.6.2474-2478.2005
|
| [16] |
Tadevosyan S, Shirvanyan A, Markosyan A, et al. (2023) Chemical composition and antibacterial activity of essential oil of Mentha arvensis L. harvested at high altitude Armenian flora. Proceedings of the YSU B: Chemical and Biological Sciences 57: 230-237. https://doi.org/10.46991/PYSU:B/2023.57.3.230
|
| [17] |
Lorenzo V, Wee S, Herrero M, et al. (1987) Operator sequences of the aerobactin operon of plasmid ColV-K30 binding the ferric uptake regulation (fur) repressor. J Bacteriol 169: 2624-2630. https://doi.org/10.1128/jb.169.6.2624-2630.1987
|
| [18] |
Maringanti S, Imlay J (1999) An intracellular iron chelator pleiotropically suppresses enzymatic and growth defects of superoxide dismutase-deficient Escherichia coli. J Bacteriol 181: 3792-3802. https://doi.org/10.1128/jb.181.12.3792-3802.1999
|
| [19] |
Hovhannisyan Z, Timotina M, Manoyan J, et al. (2022) Ribes nigrum L. extract-mediated green synthesis and antibacterial action mechanisms of silver nanoparticles. Antibiotics 11: 1415. https://doi.org/10.3390/antibiotics11101415
|
| [20] |
Sieuwerts S, De Bok FAM, Mols E, et al. (2008) A simple and fast method for determining colony forming units. Lett Appl Microbiol 47: 275-278. https://doi.org/10.1111/j.1472-765X.2008.02417.x
|
| [21] |
Vanyan L, Trchounian K (2022) HyfF subunit of hydrogenase 4 is crucial for regulating FOF1 dependent proton/potassium fluxes during fermentation of various concentrations of glucose. Jf Bioenerg Biomembr 54: 69-79. https://doi.org/10.1007/s10863-022-09930-x
|
| [22] |
Chang Y, Sun W, Murchie AIH, et al. (2023) Genome-wide identification of kanamycin B binding RNA in Escherichia coli. BMC Genomics 24: 120. https://doi.org/10.1186/s12864-023-09234-3
|
| [23] |
Mohaddab M, El Goumi Y, Gallo M, et al. (2022) Biotechnology and in vitro culture as an alternative system for secondary metabolite production. Molecules 27: 8093. https://doi.org/10.3390/molecules27228093
|
| [24] |
Chaachouay N, Zidane L (2024) Plant-derived natural products: a source for drug discovery and development. Drug Drug Candidates 3: 184-207. https://doi.org/10.3390/ddc3010011
|
| [25] |
Uritu CM, Mihai CT, Stanciu GD, et al. (2018) Medicinal plants of the family Lamiaceae in pain therapy: a review. Pain Res Manag 2018: 7801543. https://doi.org/10.1155/2018/7801543
|
| [26] |
Piran P, Kafil HS, Ghanbarzadeh S, et al. (2017) Formulation of menthol-loaded nanostructured lipid carriers to enhance its antimicrobial activity for food preservation. Adv Pharm Bull 7: 261-268. https://doi.org/10.15171/apb.2017.031
|
| [27] |
Rolfe MD, Rice CJ, Lucchini S, et al. (2012) Lag phase is a distinct growth phase that prepares bacteria for exponential growth and involves transient metal accumulation. J Bacteriol 194: 686-701. https://doi.org/10.1128/JB.06112-11
|
| [28] |
Benarroch JM, Asally M (2020) The microbiologist's guide to membrane potential dynamics. Trends Microbiol 28: 304-314. https://doi.org/10.1016/j.tim.2019.12.008
|
| [29] |
Lane N (2017) Proton gradients at the origin of life. Bioessays 39: 1600217. https://doi.org/10.1002/bies.201600217
|
| [30] |
Sapra R, Bagramyan K, Adams MW (2003) A simple energy-conserving system: proton reduction coupled to proton translocation. Proc Natl Acad Sci USA 100: 7545-7550. https://doi.org/10.1073/pnas.1331436100
|
| [31] |
Toei M, Noji H (2013) Single-molecule analysis of F0F1-ATP synthase inhibited by N,N-dicyclohexylcarbodiimide. J Biol Chem 288: 25717-25726. https://doi.org/10.1074/jbc.M113.482455
|
| [32] |
Rieger B, Arroum T, Borowski MT, et al. (2021) Mitochondrial F1 FO ATP synthase determines the local proton motive force at cristae rims. EMBO Rep 22: e52727. https://doi.org/10.15252/embr.202152727
|
Figures(3)

Silvard Tadevosyan, Naira Sahakyan. Influence of menthol on membrane-associated properties of tetracycline-resistant Escherichia coli[J]. AIMS Biophysics, 2024, 11(3): 329-339. doi: 10.3934/biophy.2024018







 DownLoad:
DownLoad: