Logistics enterprises are searching for a sustainable solution between the economy and the environment under the concept of green logistics development. Given that, this study integrates carbon emission as one of the costs into the vehicle routing problem with time window (VRPTW) and establishes a multi-center joint distribution optimization model taking into account distribution cost, carbon emission, and customer satisfaction. In the study of carbon emissions, this paper selected the vehicle load rate and vehicle distance as the main indicators. An improved ant colony algorithm is designed to solve the model by introducing the elite strategy, the saving strategy, vehicle service rules, and customer selection rules. Simulation results show that compared with the traditional ant colony optimization and genetic algorithm, the improved ant colony algorithm can effectively reduce the distribution cost and carbon emission and, improve customer satisfaction.
Citation: Xiangyang Ren, Xinxin Jiang, Liyuan Ren, Lu Meng. A multi-center joint distribution optimization model considering carbon emissions and customer satisfaction[J]. Mathematical Biosciences and Engineering, 2023, 20(1): 683-706. doi: 10.3934/mbe.2023031
Logistics enterprises are searching for a sustainable solution between the economy and the environment under the concept of green logistics development. Given that, this study integrates carbon emission as one of the costs into the vehicle routing problem with time window (VRPTW) and establishes a multi-center joint distribution optimization model taking into account distribution cost, carbon emission, and customer satisfaction. In the study of carbon emissions, this paper selected the vehicle load rate and vehicle distance as the main indicators. An improved ant colony algorithm is designed to solve the model by introducing the elite strategy, the saving strategy, vehicle service rules, and customer selection rules. Simulation results show that compared with the traditional ant colony optimization and genetic algorithm, the improved ant colony algorithm can effectively reduce the distribution cost and carbon emission and, improve customer satisfaction.

| [1] | Q. Ren, Influence of low carbon logistics industry in foreign countries on development of China's low carbon logistics industry, Environ. Sci. Manage., 43 (2018), 41-44. |
| [2] |
J. V. Hoff, M. M. B. Rasmussen, P. B. Sørensen, Barriers and opportunities in developing and implementing a Green GDP, Ecol. Econ., 181 (2021), 106905. https://doi.org/10.1016/j.ecolecon.2020.106905 doi: 10.1016/j.ecolecon.2020.106905

|
| [3] |
M. Kalantaripo, H. N. Alamdarl, Spatial effects of energy consumption and green GDP in regional agreements, Sustainability, 13 (2021), 10078. https://doi.org/10.3390/su131810078 doi: 10.3390/su131810078
|
| [4] |
H. Kaur, S. P. Singh, Heuristic modeling for sustainable procurement and logistics in a supply chain using big data, Comput. Oper. Res., 98 (2017), 301-321. https://doi.org/10.1016/j.cor.2017.05.008 doi: 10.1016/j.cor.2017.05.008
|
| [5] |
H. M. Fan, X. Yang, D. Li, Y. Li, P. Liu, J. X. Wu, Half-open multi-depot vehicle routing problem based on joint distribution mode of fresh food, Comput. Integr. Manuf. Syst., 25 (2019), 256-266. https://doi.org/10.13196/j.cims.2019.01.026 doi: 10.13196/j.cims.2019.01.026
|
| [6] |
A. I. Malik, B. S. Kim, A constrained production system involving production flexibility and carbon emissions, Mathematics, 8 (2020), 275. https://doi.org/10.3390/math8020275 doi: 10.3390/math8020275
|
| [7] |
B. Sarkar, M. Sarkar, B. Ganguly, L. E. Cárdenas-Barrón, Combined effects of carbon emission and production quality improvement for fixed lifetime products in a sustainable supply chain management, Int. J. Prod. Econ., 231 (2021), 107867. https://doi.org/10.1016/j.ijpe.2020.107867 doi: 10.1016/j.ijpe.2020.107867
|
| [8] |
T. Ghanbarpour, A. Gustafsson, How do corporate social responsibility (CSR) and innovativeness increase financial gains? A customer perspective analysis, J. Business Res., 140 (2022), 471-481. https://doi.org/10.1016/j.jbusres.2021.11.016 doi: 10.1016/j.jbusres.2021.11.016
|
| [9] |
M. Tayyab, M. S. Habib, M. S. S. Jajja, B. Sarkar, Economic assessment of a serial production system with random imperfection and shortages: A step towards sustainability, Comput. Ind. Eng., 171 (2022), 108398. https://doi.org/10.1016/j.cie.2022.108398 doi: 10.1016/j.cie.2022.108398
|
| [10] |
G. B. Dantzig, J. H. Ramser, The truck dispatching problem, Manage. Sci., 6 (1959), 80-91. https://doi.org/10.1287/mnsc.6.1.80 doi: 10.1287/mnsc.6.1.80
|
| [11] |
D. Cinar, K. Gakis, P. M. Pardalos, A 2-phase constructive algorithm for cumulative vehicle routing problems with limited duration, Expert Syst. Appl., 56 (2016), 48-58. https://doi.org/10.1016/j.eswa.2016.02.046 doi: 10.1016/j.eswa.2016.02.046
|
| [12] |
Y. Xiao, A. Konak, The heterogeneous green vehicle routing and scheduling problem with time-varying traffic congestion, Transp. Res. Part E Logist. Transp. Rev., 88 (2016), 146-166. https://doi.org/10.1016/j.tre.2016.01.011 doi: 10.1016/j.tre.2016.01.011
|
| [13] |
X. P. Deng, L. Chen, S. Tian, Research on multimodal transport path optimization with mixed time windows constraints, Int. Core J. Eng., 6 (2020), 125-129. https://doi.org/10.6919/ICJE.202003_6(3).0023 doi: 10.6919/ICJE.202003_6(3).0023
|
| [14] |
L. Y. Zhang, M. L. Tseng, C. H. Wang, C. Xiao, T. Fei, Low-carbon cold chain logistics using ribonucleic acid-ant colony optimization algorithm, J. Cleaner Prod., 233 (2019), 169-180. https://doi.org/10.1016/j.jclepro.2019.05.306 doi: 10.1016/j.jclepro.2019.05.306
|
| [15] |
T. Ning, L. An, X. Duan, Optimization of cold chain distribution path of fresh agricultural products under carbon tax mechanism: A case study in China, J. Intell. Fuzzy Syst., 40 (2021), 10549-10558. https://doi.org/10.3233/JIFS-201241 doi: 10.3233/JIFS-201241
|
| [16] |
Y. Zhang, C. Yuan, J. Wu, Vehicle routing optimization of instant distribution routing based on customer satisfaction, Information, 11 (2020), 36-36. https://doi.org/10.3390/info11010036 doi: 10.3390/info11010036
|
| [17] |
X. Li, K. Zhou, Multi-objective cold chain logistic distribution center location based on carbon emission, Environ. Sci. Pollut. Res., 28 (2021), 32396-32404. https://doi.org/10.1007/S11356-021-12992-W doi: 10.1007/S11356-021-12992-W
|
| [18] |
M. Rabbani, A. Farshbaf-Geranmayeh, N. Haghjoo, Vehicle routing problem with considering multi-middle depots for perishable food delivery, Uncertain Supply Chain Manage., 4 (2016), 171-182. https://doi.org/10.5267/j.uscm.2016.3.001 doi: 10.5267/j.uscm.2016.3.001
|
| [19] |
M. Adelzadeh, V. Mahdavi Asl, M. Koosha, A mathematical model and a solving procedure for multi-depot vehicle routing problem with fuzzy time window and heterogeneous vehicle, Int. J. Adv. Manuf. Technol., 75 (2014), 793-802. https://doi.org/10.1007/s00170-014-6141-8 doi: 10.1007/s00170-014-6141-8
|
| [20] | W. Li, X. Kou, C. Zhu, Research on optimization of joint distribution of cold chain logistics adopts carbon emission, in Journal of Physics: Conference Series, IOP Publishing, 1972 (2021), 012078. https://doi.org/10.1088/1742-6596/1972/1/012087 |
| [21] |
M. Golestani, S. H. Moosavirad, Y. Asadi, S. Biglari, A multi-objective green hub location problem with multi item-multi temperature joint distribution for perishable products in cold supply chain, Sustainable Prod. Consumption, 27 (2021), 1183-1194. https://doi.org/10.1016/j.spc.2021.02.026 doi: 10.1016/j.spc.2021.02.026
|
| [22] | D. Zhang, J. Zhang, Research on picking route optimization based on simulated annealing algorithm, in Journal of Physics: Conference Series, IOP Publishing, 1972 (2021), 012086. https://doi.org/10.1088/1742-6596/1972/1/012086 |
| [23] |
B. Liu, Logistics distribution route optimization model based on recursive fuzzy neural network algorithm, Comput. Intell. Neurosci., 2021 (2021), 3338840. https://doi.org/10.1155/2021/3338840 doi: 10.1155/2021/3338840
|
| [24] |
D. Cattaruzza, N. Absi, D. Feillet, J. González-Feliu, Vehicle routing problems for city logistics, EURO J. Transp. Logist., 6 (2017), 51-79. https://doi.org/10.1007/s13676-014-0074-0 doi: 10.1007/s13676-014-0074-0
|
| [25] |
E. B. Mariano, J. A. Gobbo Jr, F. de Castro Camioto, D. Aparecida do Nascimento Rebelatto, CO2 emissions and logistics performance: a composite index proposal, J. Cleaner Prod., 163 (2017), 166-178. https://doi.org/10.1016/j.jclepro.2016.05.084 doi: 10.1016/j.jclepro.2016.05.084
|
| [26] |
Y. Xiao, Q. Zhao, I. Kaku, Y. Xu, Development of a fuel consumption optimization model for the capacitated vehicle routing problem, Comput. Oper. Res., 39 (2012), 1419-1431. https://doi.org/10.1016/j.cor.2011.08.013 doi: 10.1016/j.cor.2011.08.013
|
| [27] | DEFRA, Guidelines for Company Reporting on Greenhouse Gas Emissions, Department for Environment, Food and Rural Affairs, 2005. |
| [28] |
X. Tian, L. Liu, S. Liu, Z. Du, M. Pang, Path planning of mobile robot based on improved ant colony algorithm for logistics, Math. Biosci. Eng., 18 (2021), 3034-3045. https://doi.org/10.3934/mbe.2021152 doi: 10.3934/mbe.2021152
|
| [29] |
Q. Yao, S. Zhu, Y. Li, Green vehicle-routing problem of fresh agricultural products considering carbon emission, Int. J. Environ. Res. Public Health, 19 (2022), 8675. https://doi.org/10.3390/ijerph19148675 doi: 10.3390/ijerph19148675
|
| [30] |
G. Qin, F. Tao, L. Li, A vehicle routing optimization problem for cold chain logistics considering customer satisfaction and carbon emissions, Int. J. Environ. Res. Public Health, 16 (2019), 576. https://doi.org/10.3390/ijerph16040576 doi: 10.3390/ijerph16040576
|
| [31] |
X. Pu, X. Lu, G. Han, An improved optimization algorithm for a multi-depot vehicle routing problem considering carbon emissions, Environ. Sci. Poll. Res., 29 (2022), 54940-54955. https://doi.org/10.1007/s11356-022-19370-0 doi: 10.1007/s11356-022-19370-0
|
| [32] |
H. Xiong, Research on cold chain logistics distribution route based on ant colony optimization algorithm, Discrete Dyn. Nat. Soc., 2021 (2021). https://doi.org/10.1155/2021/6623563 doi: 10.1155/2021/6623563
|
| [33] |
S. Yin, F. Tan, M. Yang, Summary of research on multi-objective optimization problems, Int. Core J. Eng., 7 (2021), 191-196. https://doi.org/10.6919/ICJE.202111_7(11).0032 doi: 10.6919/ICJE.202111_7(11).0032
|
| [34] |
W. Hu, K. Wu, P. P. Shum, N. I. Zheludev, C. Soci, All-optical implementation of the ant colony optimization algorithm, Sci. Rep., 6 (2016), 1-7. https://doi.org/10.1038/srep26283 doi: 10.1038/srep26283
|
| [35] |
H. Xu, P. Pu, F. Duan, Dynamic vehicle routing problems with enhanced ant colony optimization, Discrete Dyn. Nat. Soc., 2018 (2018), 1295485. https://doi.org/10.1155/2018/1295485 doi: 10.1155/2018/1295485
|
| [36] |
M. He, Z. Wei, X. Wu, Y. Peng, An adaptive variable neighborhood search ant colony algorithm for vehicle routing problem with soft time windows, IEEE Access, 9 (2021), 21258-21266. https://doi.org/10.1109/ACCESS.2021.3056067 doi: 10.1109/ACCESS.2021.3056067
|
| [37] |
D. Chen, X. M. You, S. Liu, Ant colony algorithm with Stackelberg game and multi-strategy fusion, Appl. Intell., 52 (2021), 1-23. https://doi.org/10.1007/S10489-021-02774-9 doi: 10.1007/S10489-021-02774-9
|
| [38] |
G. Qin, F. Tao, L. Li, A vehicle routing optimization problem for cold chain logistics considering customer satisfaction and carbon emissions, Int. J. Environ. Res. Public Health, 16 (2019), 576. https://doi.org/10.3390/ijerph16040576 doi: 10.3390/ijerph16040576
|
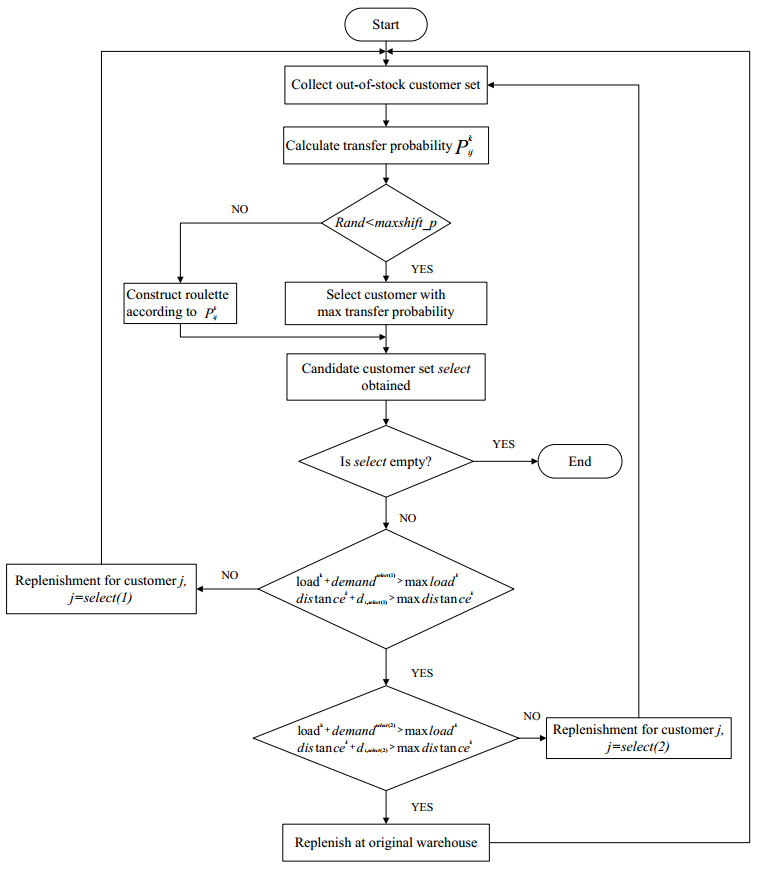
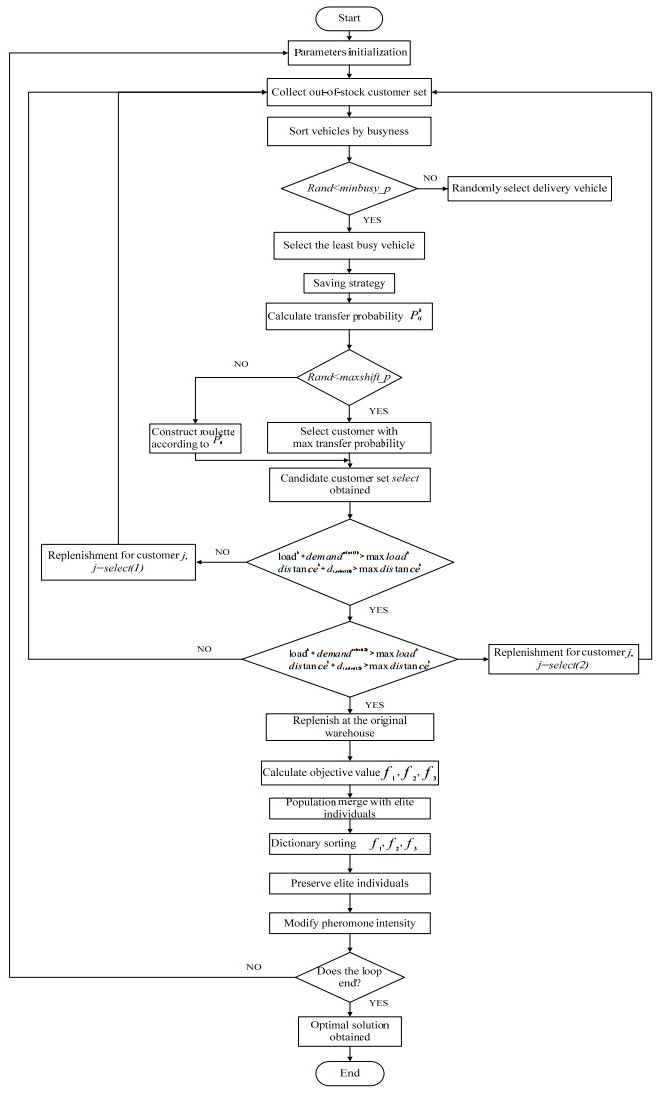
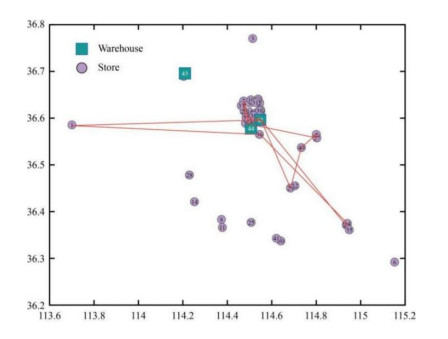
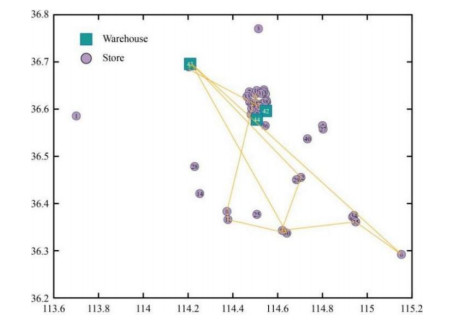
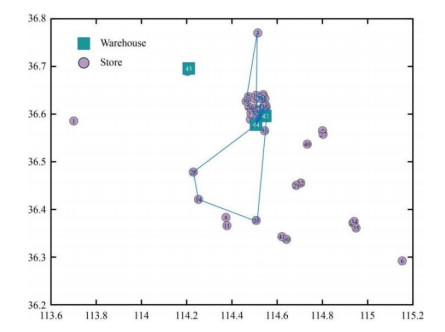
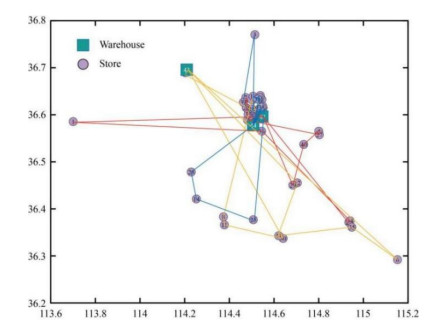
Figures(10) / Tables(5)

Xiangyang Ren, Xinxin Jiang, Liyuan Ren, Lu Meng. A multi-center joint distribution optimization model considering carbon emissions and customer satisfaction[J]. Mathematical Biosciences and Engineering, 2023, 20(1): 683-706. doi: 10.3934/mbe.2023031







 DownLoad:
DownLoad: