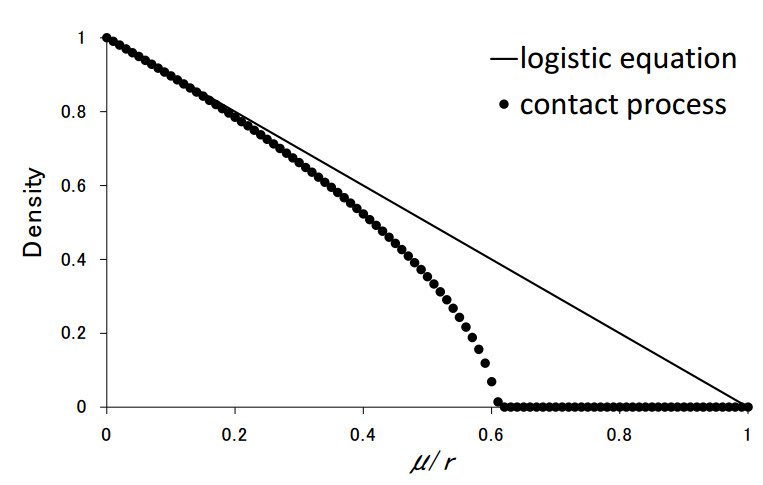
Anthropogenic modification of natural habitats is a growing threat to biodiversity and ecosystem services. The protection of biospecies has become increasingly important. Here, we pay attention to a single species as a conservation target. The species has three processes: reproduction, death and movement. Two different measures of habitat protection are introduced. One is partial protection in a single habitat (patch); the mortality rate of the species is reduced inside a rectangular area. The other is patch protection in a two-patch system, where only the mortality rate in a particular patch is reduced. For the one-patch system, we carry out computer simulations of a stochastic cellular automaton for a "contact process". Individual movements follow random walking. For the two-patch system, we assume an individual migrates into the empty cell in the destination patch. The reaction-diffusion equation (RDE) is derived, whereby the recently developed "swapping migration" is used. It is found that both measures are mostly effective for population persistence. However, comparing the results of the two measures revealed different behaviors. ⅰ) In the case of the one-patch system, the steady-state densities in protected areas are always higher than those in wild areas. However, in the two-patch system, we have found a paradox: the densities in protected areas can be lower than those in wild areas. ⅱ) In the two-patch system, we have found another paradox: the total density in both patches can be lower, even though the proportion of the protected area is larger. Both paradoxes clearly occur for the RDE with swapping migration.
Citation: Nariyuki Nakagiri, Hiroki Yokoi, Yukio Sakisaka, Kei-ichi Tainaka. Population persistence under two conservation measures: Paradox of habitat protection in a patchy environment[J]. Mathematical Biosciences and Engineering, 2022, 19(9): 9244-9257. doi: 10.3934/mbe.2022429
Anthropogenic modification of natural habitats is a growing threat to biodiversity and ecosystem services. The protection of biospecies has become increasingly important. Here, we pay attention to a single species as a conservation target. The species has three processes: reproduction, death and movement. Two different measures of habitat protection are introduced. One is partial protection in a single habitat (patch); the mortality rate of the species is reduced inside a rectangular area. The other is patch protection in a two-patch system, where only the mortality rate in a particular patch is reduced. For the one-patch system, we carry out computer simulations of a stochastic cellular automaton for a "contact process". Individual movements follow random walking. For the two-patch system, we assume an individual migrates into the empty cell in the destination patch. The reaction-diffusion equation (RDE) is derived, whereby the recently developed "swapping migration" is used. It is found that both measures are mostly effective for population persistence. However, comparing the results of the two measures revealed different behaviors. ⅰ) In the case of the one-patch system, the steady-state densities in protected areas are always higher than those in wild areas. However, in the two-patch system, we have found a paradox: the densities in protected areas can be lower than those in wild areas. ⅱ) In the two-patch system, we have found another paradox: the total density in both patches can be lower, even though the proportion of the protected area is larger. Both paradoxes clearly occur for the RDE with swapping migration.

| [1] | O. H. Frankel, M. E. Soule, Conservation and Evolution, Cambridge Univ. Press, Cambridge, 1981. |
| [2] | E. O. Wilson, The Diversity of Life, Harvard Univ. Press, Cambridge, 1992. |
| [3] |
K. L. Ryall, L. Fahrig, Habitat loss decreases predator-prey ratios in a pine-bark beetle system, Oikos, 110 (2005), 265–270. https://doi.org/10.1111/j.0030-1299.2005.13691.x doi: 10.1111/j.0030-1299.2005.13691.x

|
| [4] |
N. Nakagiri, K. Tainaka, J. Yoshimura, Bond and site percolation and habitat destruction in model ecosystems, J. Phys. Soc. Jpn., 74 (2005), 3163–3166. https://doi.org/10.1143/JPSJ.74.3163 doi: 10.1143/JPSJ.74.3163
|
| [5] |
N. Nakagiri, Y. Sakisaka, T. Togashi, S. Morita, K. Tainaka, Effects of habitat destruction in model ecosystems: Parity law depending on species richness, Ecol. Inform., 5 (2010), 241–247. https://doi.org/10.1016/j.ecoinf.2010.05.003 doi: 10.1016/j.ecoinf.2010.05.003
|
| [6] |
M. E. Gilpin, M. L. Rosenzweig, Enriched predator–prey systems: Theoretical stability, Science, 177 (1972), 902–904. https://doi.org/10.1126/science.177.4052.902 doi: 10.1126/science.177.4052.902
|
| [7] |
N. Nakagiri, K. Tainaka, T. Tao, Indirect relation between species extinction and habitat destruction, Ecol. Model., 137 (2001), 109–118. https://doi.org/10.1016/S0304-3800(00)00417-8 doi: 10.1016/S0304-3800(00)00417-8
|
| [8] |
P. P. Avelino, B. F. Oliveira, R. S. Trintin, Predominance of the weakest species in Lotka-Volterra and May-Leonard formulations of the rock-paper-scissors model, Phys. Rev. E, 100 (2019), 042209. https://link.aps.org/doi/10.1103/PhysRevE.100.042209 doi: 10.1103/PhysRevE.100.042209
|
| [9] |
K. Tainaka, N. Nakagiri, H. Yokoi, K. Sato, Multi-layered model for rock-paper-scissors game: A swarm intelligence sustains biodiversity, Ecol. Inform., 66 (2021), 101477. https://doi.org/10.1016/j.ecoinf.2021.101477 doi: 10.1016/j.ecoinf.2021.101477
|
| [10] |
N. Nakagiri, K. Tainaka, Indirect effects of habitat destruction in model ecosystems, Ecol. Model., 174 (2004), 103–114. https://doi.org/10.1016/j.ecolmodel.2003.12.047 doi: 10.1016/j.ecolmodel.2003.12.047
|
| [11] |
Y. Chen, S. J. Wright, H. C. Muller-Landau, S. P. Hubbell, Y. Wang, S. Yu, Positive effects of neighborhood complementarity on tree growth in a Neotropical forest, Ecology, 97 (2016), 776–785. https://doi.org/10.1890/15-0625.1 doi: 10.1890/15-0625.1
|
| [12] |
M. Perc, Does strong heterogeneity promote cooperation by group interactions?, New J. Phys., 13 (2011), 123027. https://doi.org/10.1088/1367-2630/13/12/123027 doi: 10.1088/1367-2630/13/12/123027
|
| [13] |
G. Szabó, G. Fáth, Evolutionary games on graphs, Phys. Rep., 446 (2007), 97–216. https://doi.org/10.1016/j.physrep.2007.04.004 doi: 10.1016/j.physrep.2007.04.004
|
| [14] |
A. Szolnoki, M. Perc, Evolutionary dynamics of cooperation in neutral populations, New J. Phy., 20 (2018), 013031. https://doi.org/10.1088/1367-2630/aa9fd2 doi: 10.1088/1367-2630/aa9fd2
|
| [15] |
K. Tainaka, Lattice model for the Lotka-Volterra system, J. Phys. Soc. Jpn., 57 (1988), 2588–2590. https://doi.org/10.1143/JPSJ.57.2588 doi: 10.1143/JPSJ.57.2588
|
| [16] |
K. Tainaka, Stationary pattern of vortices or strings in biological systems: lattice version of the Lotka-Volterra model, Phys. Rev. Lett., 63 (1989), 2688–2691. https://doi.org/10.1103/PhysRevLett.63.2688 doi: 10.1103/PhysRevLett.63.2688
|
| [17] |
T. E. Harris, Contact interaction on a lattice, Ann. Prob., 2 (1974), 969–988. https://doi.org/10.1214/aop/1176996493 doi: 10.1214/aop/1176996493
|
| [18] |
J. Tubay, H. Ito, T. Uehara, S. Kakishima, S. Morita, T. Togashi, et al., The paradox of enrichment in phytoplankton by induced competitive interactions, Sci. Rep., 3 (2013), 2835. https://doi.org/10.1038/srep02835 doi: 10.1038/srep02835
|
| [19] |
N. Nakagiri, K. Sato, Y. Sakisaka, K. Tainaka, Serious role of non-quarantined COVID-19 patients for random walk simulations, Sci. Rep., 12 (2022), 738. https://doi.org/10.1038/s41598-021-04629-2 doi: 10.1038/s41598-021-04629-2
|
| [20] |
T. Tahara, M. K. A. Gavina, T. Kawano, J. M. Tubay, J. F. Rabajante, H. Ito, et al., Asymptotic stability of a modified Lotka-Volterra model with small immigrations, Sci. Rep., 8 (2018), 7029. https://doi.org/10.1038/s41598-018-25436-2 doi: 10.1038/s41598-018-25436-2
|
| [21] | I. Hanski, Metapopulation Ecology, Oxford University Press, New York, 1999. |
| [22] | I. Hanski, M. E. Gilpin, Metapopulation Biology: Ecology, Genetics, and Evolution, Academic Press, San Diego, 1997. |
| [23] |
S. A. Levin, Dispersion and population interactions, Am. Nat., 108 (1974), 207–228. https://doi.org/10.1086/282900 doi: 10.1086/282900
|
| [24] |
K. M. A. Kabir, J. Tanimoto, Analysis of epidemic outbreaks in two-layer networks with different structures for information spreading and disease diffusion, Commun. Nonlinear Sci. Numer. Simul., 72 (2019), 565–574. https://doi.org/10.1016/j.cnsns.2019.01.020 doi: 10.1016/j.cnsns.2019.01.020
|
| [25] |
K. M. A. Kabir, J. Tanimoto, Evolutionary vaccination game approach in metapopulation migration model with information spreading on different graphs, Chaos Solitons Fractals, 120 (2019), 41–55. https://doi.org/10.1016/j.chaos.2019.01.013 doi: 10.1016/j.chaos.2019.01.013
|
| [26] |
T. Nagatani, G. Ichinose, K. Tainaka, Heterogeneous network promotes species coexistence: metapopulation model for rock-paper-scissors game, Sci. Rep., 8 (2018), 7094. https://doi.org/10.1038/s41598-018-25353-4 doi: 10.1038/s41598-018-25353-4
|
| [27] |
A. Sadykov, K. D. Farnsworth, Model of two competing populations in two habitats with migration: Application to optimal marine protected area size, Theor. Popul. Biol., 142 (2021), 114–122. https://doi.org/10.1016/j.tpb.2021.10.002 doi: 10.1016/j.tpb.2021.10.002
|
| [28] |
H. Yokoi, K. Tainaka, K. Sato, Metapopulation model for a prey-predator system: Nonlinear migration due to the finite capacities of patches, J. Theor. Biol., 477 (2019), 24–35. https://doi.org/10.1016/j.jtbi.2019.05.021 doi: 10.1016/j.jtbi.2019.05.021
|
| [29] |
H. Yokoi, K. Tainaka, N. Nakagiri, K. Sato, Self-organized habitat segregation in an ambush-predator system: Nonlinear migration of prey between two patches with finite capacities, Ecol. Inform., 55 (2020), 101022. https://doi.org/10.1016/j.ecoinf.2019.101022 doi: 10.1016/j.ecoinf.2019.101022
|
| [30] | T. M. Liggett, Interacting Particle Systems, Springer, New York, 1985. https://doi.org/10.1007/978-1-4613-8542-4 |
| [31] | J. Marro, R. Dickman, Nonequilibrium Phase Transition in Lattice Models, Cambridge University Press, Cambridge, 1999. https://doi.org/10.1017/CBO9780511524288 |
| [32] | N. Konno, Phase Transition of Interacting Particle Systems, World Scientific, Singapore, 1991. |
| [33] |
M. Katori, N. Konno, Upper bounds for survival probability of the contact process, J. Stat. Phys., 63 (1991), 115–130. https://doi.org/10.1007/BF01026595 doi: 10.1007/BF01026595
|
| [34] |
W. Cota, A. S. Mata, S. C. Ferreira, Robustness and fragility of the susceptible-infected-susceptible epidemic models on complex networks, Phys. Rev. E, 98 (2018), 012310. https://doi.org/10.1103/PhysRevE.98.012310 doi: 10.1103/PhysRevE.98.012310
|
| [35] |
T. Gross, C. J. D'Lima, B. Blasius, Epidemic dynamics on an adaptive network, Phys. Rev. Lett., 96 (2006), 208701. https://doi.org/10.1103/PhysRevLett.96.208701 doi: 10.1103/PhysRevLett.96.208701
|
| [36] |
J. Peterson, The contact process on the complete graph with random vertex-dependent infection rates, Stoch. Proc. Their Appl., 121 (2011), 609–629. https://doi.org/10.1016/j.spa.2010.11.003 doi: 10.1016/j.spa.2010.11.003
|
| [37] |
Y. Harada, H. Ezoe, Y. Iwasa, H. Matsuda, K. Sato, Population persistence and spatially limited social interaction, Theor. Popul. Biol., 48 (1995), 65–91. https://doi.org/10.1006/tpbi.1995.1022 doi: 10.1006/tpbi.1995.1022
|
| [38] |
K. Tainaka, N. Nakagiri, Segregation in an interacting particle system, Phys. Lett. A, 271 (2000), 92–99. https://doi.org/10.1016/S0375-9601(00)00302-9 doi: 10.1016/S0375-9601(00)00302-9
|
| [39] |
J. M. Tubay, J. Yoshimura, Resistance of a terrestrial plant community to local microhabitat changes, Ecol. Evol., 8 (2018), 5101–5110. https://doi.org/10.1002/ece3.4093 doi: 10.1002/ece3.4093
|
| [40] |
X. Zou, K. Wang, A robustness analysis of biological population models with protection zone, Appl. Math. Model., 35 (2011), 5553–5563. https://doi.org/10.1016/j.apm.2011.05.020 doi: 10.1016/j.apm.2011.05.020
|
| [41] |
F. Wei, C. Wang, Survival analysis of a single-species population model with fluctuations and migrations between patches, Appl. Math. Model., 81 (2020), 113–127. https://doi.org/10.1016/j.apm.2019.12.023 doi: 10.1016/j.apm.2019.12.023
|







Figures(8) / Tables(1)

Nariyuki Nakagiri, Hiroki Yokoi, Yukio Sakisaka, Kei-ichi Tainaka. Population persistence under two conservation measures: Paradox of habitat protection in a patchy environment[J]. Mathematical Biosciences and Engineering, 2022, 19(9): 9244-9257. doi: 10.3934/mbe.2022429







 DownLoad:
DownLoad: