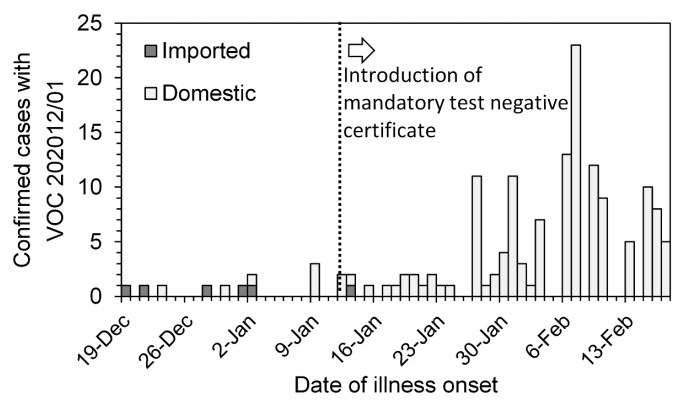
Coronavirus disease 2019 (COVID-19), caused by severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2), has rapidly spread across the globe. The variant of concern (VOC) 202012/01 (B.1.1.7, also known as the alpha variant) bearing the N501Y mutation emerged in late 2020. VOC 202012/01 was more transmissible than existing SARS-CoV-2 variants and swiftly became dominant in many regions. More than 150 cases of VOC 202012/01 were reported in Japan by 26 February 2021. During the very early stage of introduction, only a subset arose from domestic transmission. If the reproduction number R (i.e., the average number of secondary transmission events caused by a single primary case) is greater than 1, the corresponding proportion should converge to 1 in a short period of time, and thus it is critical to understand the transmissibility of VOC 202012/01 based on travel history information. The present study aimed to estimate R of VOC 202012/01 using overseas travel history information. A mathematical model was developed to capture the relationship between travel history and R. We obtained travel history data for each confirmed case of VOC 202012/01 infection from 26 December 2020 to 26 February 2021. Maximum likelihood estimation was used to estimate R, accounting for right censoring during real-time estimation. In the baseline scenario, R was estimated at 2.11 (95% confidence interval: 1.63, 2.94). By 26 February 2021, an average of nine generations had elapsed since the first imported case. If the generation time of VOC 202012/01 was assumed to be longer, R was increased, consistent with estimates of R from case data. The estimated R of VOC 202012/01 in Japan exceeded 1 on 26 February 2021, suggesting that domestic transmission events caused a major epidemic. Moreover, because our estimate of R was dependent on generation time and ascertainment biases, continuous monitoring of contact tracing data is crucial to decipher the mechanisms of increased VOC 202012/01 transmissibility.
Citation: Taishi Kayano, Hiroshi Nishiura. Estimating the transmissibility of SARS-CoV-2 VOC 202012/01 in Japan using travel history information[J]. Mathematical Biosciences and Engineering, 2022, 19(3): 2750-2761. doi: 10.3934/mbe.2022125
Coronavirus disease 2019 (COVID-19), caused by severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2), has rapidly spread across the globe. The variant of concern (VOC) 202012/01 (B.1.1.7, also known as the alpha variant) bearing the N501Y mutation emerged in late 2020. VOC 202012/01 was more transmissible than existing SARS-CoV-2 variants and swiftly became dominant in many regions. More than 150 cases of VOC 202012/01 were reported in Japan by 26 February 2021. During the very early stage of introduction, only a subset arose from domestic transmission. If the reproduction number R (i.e., the average number of secondary transmission events caused by a single primary case) is greater than 1, the corresponding proportion should converge to 1 in a short period of time, and thus it is critical to understand the transmissibility of VOC 202012/01 based on travel history information. The present study aimed to estimate R of VOC 202012/01 using overseas travel history information. A mathematical model was developed to capture the relationship between travel history and R. We obtained travel history data for each confirmed case of VOC 202012/01 infection from 26 December 2020 to 26 February 2021. Maximum likelihood estimation was used to estimate R, accounting for right censoring during real-time estimation. In the baseline scenario, R was estimated at 2.11 (95% confidence interval: 1.63, 2.94). By 26 February 2021, an average of nine generations had elapsed since the first imported case. If the generation time of VOC 202012/01 was assumed to be longer, R was increased, consistent with estimates of R from case data. The estimated R of VOC 202012/01 in Japan exceeded 1 on 26 February 2021, suggesting that domestic transmission events caused a major epidemic. Moreover, because our estimate of R was dependent on generation time and ascertainment biases, continuous monitoring of contact tracing data is crucial to decipher the mechanisms of increased VOC 202012/01 transmissibility.

| [1] | Investigation of SARS-CoV-2 Variants of Concern in England, Public Health England, 2021. Available from: moz-extension://73815545-76cf-4708-8876-11f04da78d19/enhanced-reader.html?openApp&pdf=https%3A%2F%2Fassets.publishing.service.gov.uk%2Fgovernment%2Fuploads%2Fsystem%2Fuploads%2Fattachment_data%2Ffile%2F961299%2FVariants_of_Concern_VOC_Technical_Briefing_6. |
| [2] |
K. Kupferschmidt, Danish scientists see tough times ahead as variant rises, Science, 371 (2021), 549–550. doi: 10.1126/science.371.6529.549. doi: 10.1126/science.371.6529.549

|
| [3] | SARS-CoV-2 – Increased Circulation of Variants of Concern and Vaccine Rollout in the EU/EEA – 14th Update, European Centre for Disease Prevention and Control, 2021. Available from: https://www.ecdc.europa.eu/en/covid-19/timeline-ecdc-response. |
| [4] |
T. Alpert, A. F. Brito, E. Lasek-Nesselquist, J. Rothman, A. L. Valesano, M. J. MacKay, et al., Early introductions and community transmission of SARS-CoV-2 variant B.1.1.7 in the United States, Cell, 184 (2021), 2595–2604. doi: 10.1016/j.cell.2021.03.061. doi: 10.1016/j.cell.2021.03.061
|
| [5] |
R. Wölfel, V. M. Corman, W. Guggemos, M. Seilmaier, S. Zange, M. A. Müller, et al., Virological assessment of hospitalized patients with COVID-2019, Nature, 581 (2020), 465–469. doi: 10.1038/s41586-020-2196-x. doi: 10.1038/s41586-020-2196-x
|
| [6] |
S. M. Jung, A. R. Akhmetzhanov, K. Hayashi, N. M. Linton, Y. Yang, B. Yuan, et al., Real-time estimation of the risk of death from novel coronavirus (COVID-19) infection: Inference using exported cases, J. Clin. Med., 9 (2020), 523. doi: 10.3390/jcm9020523. doi: 10.3390/jcm9020523
|
| [7] |
S. Zhao, Q. Lin, J. Ran, S. S. Musa, G. Yang, W. Wang, et al., Preliminary estimation of the basic reproduction number of novel coronavirus (2019-nCoV) in China, from 2019 to 2020: A data-driven analysis in the early phase of the outbreak, Int. J. Infect. Dis., 92 (2020), 214–217. doi: 10.1016/j.ijid.2020.01.050. doi: 10.1016/j.ijid.2020.01.050
|
| [8] | Tracking Public Health and Social Measures, World Health Organization, 2019. Available from: https://www.who.int/emergencies/diseases/novel-coronavirus-2019/phsm. |
| [9] |
N. G. Davies, S. Abbott, R. C. Barnard, C. I. Jarvis, A. J. Kucharski, J. D. Munday, et al., Estimated transmissibility and impact of SARS-CoV-2 lineage B.1.1.7 in England, Science, 372 (2021), eabg3055. doi: 10.1126/science.abg3055. doi: 10.1126/science.abg3055
|
| [10] |
S. Zhao, J. Lou, L. Cao, H. Zheng, M. K. C. Chong, Z. Chen, et al., Quantifying the transmission advantage associated with N501Y substitution of SARS-CoV-2 in the UK: An early data-driven analysis, J. Travel. Med., 28 (2021), taab011. doi: 10.1093/jtm/taab011. doi: 10.1093/jtm/taab011
|
| [11] |
K. Leung, M. H. Shum, G. M. Leung, T. T. Lam, J. T. Wu, Early transmissibility assessment of the N501Y mutant strains of SARS-CoV-2 in the United Kingdom, October to November 2020, Euro. Surveill., 26 (2020), 2002106. doi: 10.2807/1560-7917.ES.2020.26.1.2002106. doi: 10.2807/1560-7917.ES.2020.26.1.2002106
|
| [12] | NERVTAG Paper on COVID-19 Variant of Concern B.1.1.7, New and Emerging Respiratory Virus Threats Advisory Group, 2021. Available from: https://www.gov.uk/government/publications/nervtag-paper-on-covid-19-variant-of-concern-b117. |
| [13] | N. G. Davies, C. I. Jarvis, CMMID COVID-19 Working Group, W. J. Edmunds, N. P. Jewell, K. Diaz-Ordaz, et al., Increased mortality in community-tested cases of SARS-CoV-2 lineage B.1.1.7, Nature, 593 (2021), 270–274. doi: 0.1038/s41586-021-03426-1. |
| [14] |
R. Challen, E. Brooks-Pollock, J. M. Read, L. Dyson, K. Tsaneva-Atanasova, L. Danon, Risk of mortality in patients infected with SARS-CoV-2 variant of concern 202012/1: Matched cohort study, BMJ, 372 (2021), n579. doi: 10.1136/bmj.n579. doi: 10.1136/bmj.n579
|
| [15] | Information About New Response for Quarantine in Japan, Ministry of Health, Labour, and Welfare of Japan, 2021. Available from: https://www.mhlw.go.jp/stf/seisakunitsuite/bunya/0000121431_00209.html. |
| [16] |
H. Nishiura, K. Mizumoto, K. Ejima, How to interpret the transmissibility of novel influenza A(H7N9): An analysis of initial epidemiological data of human cases from China, Theor. Biol. Med. Model., 10 (2013), 1–9. doi: 10.1186/1742-4682-10-30. doi: 10.1186/1742-4682-10-30
|
| [17] |
S. Cauchemez, S. Epperson, M. Biggerstaff, D. Swerdlow, L. Finelli, N. M. Ferguson, Using routine surveillance data to estimate the epidemic potential of emerging zoonoses: Application to the emergence of US swine origin influenza A H3N2v virus, PLoS Med., 10 (2013): e1001399. doi: 10.1371/journal.pmed.1001399. doi: 10.1371/journal.pmed.1001399
|
| [18] |
H. Murayama, T. Kayano, H. Nishiura, Estimating COVID-19 cases infected with the variant alpha (VOC 202012/01): An analysis of screening data in Tokyo, January–March 2021, Theor. Biol. Med. Model., 18 (2021), 13. doi: 10.1186/s12976-021-00146-x. doi: 10.1186/s12976-021-00146-x
|
| [19] | Information About the Emergences of COVID-19 Cases Associated with Variant Strains, Ministry of Health, Labour, and Welfare of Japan, 2021. Available from: https://www.mhlw.go.jp/stf/newpage_17211.html. |
| [20] |
H. Nishiura, N. M. Linton, A. R. Akhmetzhanov, Serial interval of novel coronavirus (COVID-19) infections, Int. J. Infect. Dis., 93 (2020), 284–286. doi: 10.1016/j.ijid.2020.02.060. doi: 10.1016/j.ijid.2020.02.060
|
| [21] | Confirmed Number of Cases with Variant in Each Prefecture in Japan (Genome Sequencing), Ministry of Health, Labour, and Welfare of Japan, 2021. Available from: https://www.mhlw.go.jp/content/10906000/000751599.pdf. |
| [22] |
L. M. Kucirka, S. A. Lauer, O. Laeyendecker, D. Boon, J. Lessler, Variation in false-negative rate of reverse transcriptase polymerase chain reaction-based SARS-CoV-2 tests by time since exposure, Ann. Intern. Med., 173 (2020), 262–267. doi: 10.7326/M20-1495. doi: 10.7326/M20-1495
|
| [23] | E. Volz, S. Mishra, M. Chand, J. C. Barrett, R. Johnson, L. Geidelberg, et al., Transmission of SARS-CoV-2 lineage B.1.1.7 in England: Insights from linking epidemiological and genetic data, MedRxiv, 2020.12.30.20249034 (2020). doi: https://doi.org/10.1101/2020.12.30.20249034. |
| [24] | Japan Adds 13 Nations for Extra Quarantine Steps over COVID-19 Variants, The Japan Times, 2021. Available from: https://www.japantimes.co.jp/news/2021/03/03/national/new-quarantine-measures-variants/. |


Figures(3)

Taishi Kayano, Hiroshi Nishiura. Estimating the transmissibility of SARS-CoV-2 VOC 202012/01 in Japan using travel history information[J]. Mathematical Biosciences and Engineering, 2022, 19(3): 2750-2761. doi: 10.3934/mbe.2022125







 DownLoad:
DownLoad: