Citation: Xiaoqin Wang, Yiping Tan, Yongli Cai, Kaifa Wang, Weiming Wang. Dynamics of a stochastic HBV infection model with cell-to-cell transmission and immune response[J]. Mathematical Biosciences and Engineering, 2021, 18(1): 616-642. doi: 10.3934/mbe.2021034

| [1] | World Health Organization, Hepatitis B, Accessed 12 Nov. 2020. Available from: https://www.who.int/health-topics/hepatitis#tab=tab_1. |
| [2] |
M. Nowak, R. M. May, Population dynamics of immune responses to persistent viruses, Science, 272 (1996), 74–79. doi: 10.1126/science.272.5258.74

|
| [3] |
R. A. Arnaout, M. A. Nowak, Competitive coexistence in antiviral immunity, J. Theor. Biol., 204 (2000), 431–441. doi: 10.1006/jtbi.2000.2027
|
| [4] |
R. V. Culshaw, S. Ruan, R. J. Spiteri, Optimal HIV treatment by maximising immune response, J. Math. Biol., 48 (2004), 545–562. doi: 10.1007/s00285-003-0245-3
|
| [5] | D. Bertacchi, F. Zucca, S. Foresti, D. Mangioni, A. Gori, Combination versus sequential monotherapy in chronic HBV infection: a mathematical approach, Math. Med. Biol., 32 (2015), 383–403. |
| [6] | A. U. Neumann, Hepatitis B viral kinetics: A dynamic puzzle still to be resolved, Hepatology, 42 (2005), 249–254. |
| [7] | M. Nowak, R. M. May, Virus Dynamics: Mathematical Principles of Immunology and Virology: Mathematical Principles of Immunology and Virology, Oxford University Press, UK, 2000. |
| [8] |
J. Li, K. Men, Y. Yang, D. Li, Dynamical analysis on a chronic hepatitis C virus infection model with immune response, J. Theor. Biol., 365 (2015), 337–346. doi: 10.1016/j.jtbi.2014.10.039
|
| [9] |
M. Li, J. Zu, The review of differential equation models of HBV infection dynamics, J. Virol. Meth., 266 (2019), 103–113. doi: 10.1016/j.jviromet.2019.01.014
|
| [10] | A. Mojaver, H. Kheiri, Dynamical analysis of a class of hepatitis C virus infection models with application of optimal control, Inter. J. Biomath., 9 (2016), 1650038. |
| [11] | W. Mothes, N. M. Sherer, J. Jin, P. Zhong, Virus cell-to-cell transmission, J. Virol., 84 (2010), 8360–8368. |
| [12] |
S. Pan, S. P. Chakrabarty, Threshold dynamics of HCV model with cell-to-cell transmission and a non-cytolytic cure in the presence of humoral immunity, Commun. Nonlinear Sci., 61 (2018), 180–197. doi: 10.1016/j.cnsns.2018.02.010
|
| [13] | K. Razali, H. Thein, J. Bell, M. Cooper-Stanbury, K. Dolan, G. Dore, et al., Modelling the hepatitis C virus epidemic in Australia, Drug Alcohol Depen., 91 (2007), 228–235. |
| [14] | B. Roe, W. W. Hall, Cellular and molecular interactions in coinfection with hepatitis C virus and human immunodeficiency virus, Expert Rev. Mol. Med., 10 (2008), e30. |
| [15] |
F. Zhang, J. Li, C. Zheng, L. Wang, Dynamics of an HBV/HCV infection model with intracellular delay and cell proliferation, Commun. Nonlinear Sci., 42 (2017), 464–476. doi: 10.1016/j.cnsns.2016.06.009
|
| [16] | P. Zhong, L. M. Agosto, J. B. Munro, W. Mothes, Cell-to-cell transmission of viruses, Curr. Opin. Virol., 3 (2013), 44–50. |
| [17] | M. Sourisseau, N. Sol-Foulon, F. Porrot, F. Blanchet, O. Schwartz, Inefficient human immunodeficiency virus replication in mobile lymphocytes, J. Virol., 81 (2006), 1000–1012. |
| [18] |
B. Monel, E. Beaumont, D. Vendrame, O. Schwartz, D. Brand, Hiv cell-to-cell transmission requires the production of infectious virus particles and does not proceed through env-mediated fusion pores, J. Virology, 86 (2012), 3924–3933. doi: 10.1128/JVI.06478-11
|
| [19] |
Y. Yang, Q. Han, Z. Hou, C. Zhang, Z. Tian, J. Zhang, Exosomes mediate hepatitis B virus (HBV) transmission and NK-cell dysfunction, Cell. Mol. Immunol., 14 (2017), 465–475. doi: 10.1038/cmi.2016.24
|
| [20] | G. Bocharov, B. Ludewig, A. Bertoletti, P. Klenerman, T. Junt, P. Krebs, et al., Underwhelming the immune response: effect of slow virus growth on CD8+-T-lymphocyte responses, J. Virol., 78 (2004), 2247–2254. |
| [21] |
S. Cai, Y. Cai, X. Mao, A stochastic differential equation SIS epidemic model with two independent brownian motions, J. Math. Anal. Appl., 474 (2019), 1536–1550. doi: 10.1016/j.jmaa.2019.02.039
|
| [22] |
Y. Cai, J. Jiao, Z. Gui, Y. Liu, W. M. Wang, Environmental variability in a stochastic epidemic model, Appl. Math. Comp., 329 (2018), 210–226. doi: 10.1016/j.amc.2018.02.009
|
| [23] |
Y. Cai, Y. Kang, M. Banerjee, W.M. Wang, A stochastic SIRS epidemic model with infectious force under intervention strategies, J. Differ. Equations, 259 (2015), 7463–7502. doi: 10.1016/j.jde.2015.08.024
|
| [24] |
Y. Cai, Y. Kang, W. M. Wang, A stochastic SIRS epidemic model with nonlinear incidence rate, Appl. Math. Comp., 305 (2017), 221–240. doi: 10.1016/j.amc.2017.02.003
|
| [25] | C. Ji, D. Jiang, Dynamics of an HIV-1 infection model with cell-mediated immune response and stochastic perturbation, Inter. J. Biomath., 5 (2012), 1250039. |
| [26] |
T. Luzyanina, G. Bocharov, Stochastic modeling of the impact of random forcing on persistent hepatitis B virus infection, Math. Comp. Simul., 96 (2014), 54–65. doi: 10.1016/j.matcom.2011.10.002
|
| [27] | F. A. Rihan, H. J. Alsakaji, C. Rajivganthi, Stochastic SIRC epidemic model with time-delay for covid-19, Adv. Differ. Equ., 2020 (2020), 502. |
| [28] | Y. Wang, D. Jiang, Stationary distribution and extinction of a stochastic viral infection model, Discrete Dyn. Nat. Soc., 2017 (2017), 1–13. |
| [29] |
Y. Wang, D. Jiang, A. Alsaedi, T. Hayat, Modelling a stochastic HIV model with logistic target cell growth and nonlinear immune response function, Phys. A, 501 (2018), 276–292. doi: 10.1016/j.physa.2018.02.040
|
| [30] |
Y. Wang, D. Jiang, T. Hayat, B. Ahmad, A stochastic HIV infection model with T-cell proliferation and CTL immune response, Appl. Math. Comp., 315 (2017), 477–493. doi: 10.1016/j.amc.2017.07.062
|
| [31] |
Y. Yuan, L. J. S. Allen, Stochastic models for virus and immune system dynamics, Math. Biosci., 234 (2011), 84–94. doi: 10.1016/j.mbs.2011.08.007
|
| [32] |
F. Xie, M. Shan, X. Lian, W. M. Wang, Periodic solution of a stochastic HBV infection model with logistic hepatocyte growth, Appl. Math. Comp., 293 (2017), 630–641. doi: 10.1016/j.amc.2016.06.028
|
| [33] |
T. G. Kurtz, Solutions of ordinary differential equations as limits of pure jump Markov processes, J. Appl. Probab., 7 (1970), 49–58. doi: 10.2307/3212147
|
| [34] |
T. G. Kurtz, Limit theorems for sequences of jump Markov processes approximating ordinary differential processes, J. Appl. Probab., 8 (1971), 344–356. doi: 10.2307/3211904
|
| [35] |
P. Van den Driessche, J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002), 29–48. doi: 10.1016/S0025-5564(02)00108-6
|
| [36] | X. Mao, Stochastic Differential Equations and Applications, Elsevier, 2007. |
| [37] |
D. J. Higham, An algorithmic introduction to numerical simulations of stochastic differentila equations, SIAM Rev., 43 (2001), 525–546. doi: 10.1137/S0036144500378302
|
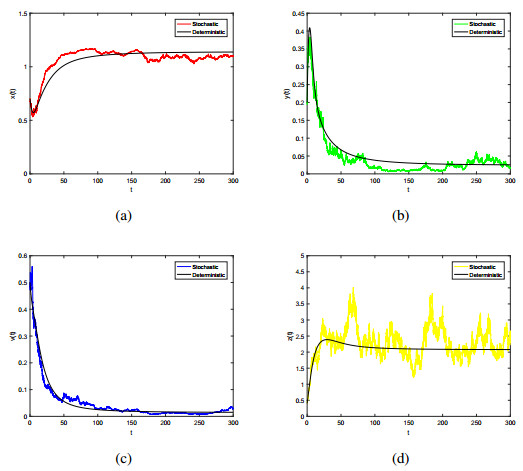
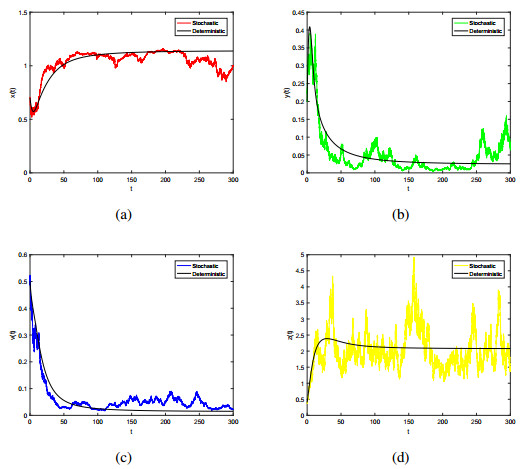





Figures(7) / Tables(1)

Xiaoqin Wang, Yiping Tan, Yongli Cai, Kaifa Wang, Weiming Wang. Dynamics of a stochastic HBV infection model with cell-to-cell transmission and immune response[J]. Mathematical Biosciences and Engineering, 2021, 18(1): 616-642. doi: 10.3934/mbe.2021034







 DownLoad:
DownLoad: