Citation: Chunxiang Fan, Zouqin Huang, Binbin Chen, Baojin Chen, Qi Wang, Weidong Liu, Donghai Yu. Comprehensive analysis of key lncRNAs in ischemic stroke[J]. Mathematical Biosciences and Engineering, 2020, 17(2): 1318-1328. doi: 10.3934/mbe.2020066

| [1] | A. Arboixand and J. Alio, Cardioembolic Stroke: Clinical Features, Specific Cardiac Disorders and Prognosis, Curr. Cardiol. Rev., 6 (2010), 150-161. |
| [2] | S. Sabour, The Diagnostic Value of Serum miRNA-221-3p, miRNA-382-5p, and miRNA-4271 in Ischemic Stroke: Methodological Issue to Avoid Misinterpretation, J. Stroke Cerebrovasc. Dis., 26 (2017), 1161. |
| [3] | Z. Wang, Y. Yuan, Z. Zhang, et al., Inhibition of miRNA-27b enhances neurogenesis via AMPK activation in a mouse ischemic stroke model, FEBS Open Bio., 9 (2019), 859-869. |
| [4] | Z. Wu, P. Wu, X. Zuo, et al., Erratum to: LncRNA-N1LR Enhances Neuroprotection Against Ischemic Stroke Probably by Inhibiting p53 Phosphorylation, Mol. Neurobiol., 54 (2017), 7670-7685. |
| [5] | J. Wang, B. Cao, D. Han, et al., Long Non-coding RNA H19 Induces Cerebral Ischemia Reperfusion Injury via Activation of Autophagy, Aging Dis., 8 (2017), 71-84. |
| [6] | J. A. Saugstad, Non-Coding RNAs in Stroke and Neuroprotection, Front. Neurol., 6 (2015), 50. |
| [7] | T. L. Barr, R. L. Vangilder, S. L. Rellick, et al., A Genomic Profile of the Immune Response to Stroke With Implications for Stroke Recovery, Biol. Res. Nurs., 17 (2015), 248-256. |
| [8] | X. Wan, W. Huang, S. Yang, et al., Identification of androgen-responsive lncRNAs as diagnostic and prognostic markers for prostate cancer, Oncotarget, 7 (2016), 60503-60518. |
| [9] | H. Lin, M. Jiang, L. Liu, et al., The long noncoding RNA Lnczc3h7a promotes a TRIM25-mediated RIG-I antiviral innate immune response, Nat. Immunol., 2019 (2019), 1. |
| [10] | C. H. Liand Y. Chen, Insight Into the Role of Long Noncoding RNA in Cancer Development and Progression, Int. Rev. Cell Mol. Biol., 326 (2016), 33-65. |
| [11] | H. Yang, X. Xi, B. Zhao, et al., KLF4 protects brain microvascular endothelial cells from ischemic stroke induced apoptosis by transcriptionally activating MALAT1, Biochem. Biophys. Res. Commun., 495 (2018), 2376-2382. |
| [12] | X. Zhang, X. L. Zhu, B. Y. Ji, et al., LncRNA-1810034E14Rik reduces microglia activation in experimental ischemic stroke, J. Neuroinflammation, 16 (2019), 75. |
| [13] | D. Wu, Y. C. G. Lee, H. C. Liu, et al., Identification of TLR downstream pathways in stroke patients. Clin. Biochem., 46 (2013):1058-1064. |
| [14] | X. Wang, S. Cheng, V. H. Brophy, et al., A Meta-Analysis of Candidate Gene Polymorphisms and Ischemic Stroke in 6 Study Populations: Association of Lymphotoxin-Alpha in Nonhypertensive Patients. Stroke, 40 (2009):683-695. |
| [15] | C. D. Anderson, A. Biffi, M. A. Nalls, et al., Common Variants Within Oxidative Phosphorylation Genes Influence Risk of Ischemic Stroke and Intracerebral Hemorrhage. Stroke, 44 (2013):612-619. |
| [16] | T. Liu, Z. Han, H. Li, et al., LncRNA DLEU1 contributes to colorectal cancer progression via activation of KPNA3, Mol. Cancer, 17 (2018), 118. |
| [17] | J. Wang, D. Pappas, P. L. De Jager, et al., Modeling the Cumulative Genetic Risk for Multiple Sclerosis from Genome-Wide Association Data, Genome Med., 3 (2011), 3. |
| [18] | T. Kim, S. J. Choi, Y. H. Lee, et al., Gene expression profile predicting the response to anti-TNF treatment in patients with rheumatoid arthritis; analysis of GEO datasets, Jt. Bone Spine, 81 (2014), 325-330. |
| [19] | G. W. E. Santen, M. Kriek and H. Van Attikum, SWI/SNF complex in disorder: SWItching from malignancies to intellectual disability, Epigenetics, 7 (2012), 1219-1224. |
| [20] | A. Saschen, M. R. Aure, L. Leibovich, et al., LIMT is a novel metastasis inhibiting lncRNA suppressed by EGF and downregulated in aggressive breast cancer, Embo Mol. Med., 8 (2016), 1052-1064. |
| [21] | J. D. J. Labonne, T. Graves, Y. Shen, et al., A microdeletion at Xq22.2 implicates a glycine receptor GLRA4 involved in intellectual disability, behavioral problems and craniofacial anomalies, BMC Neurol., 16 (2016), 132. |
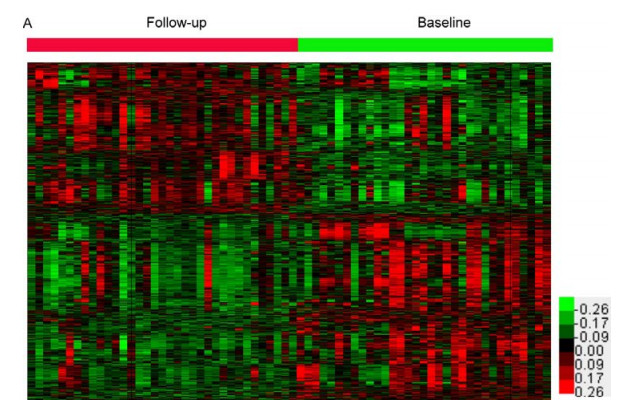
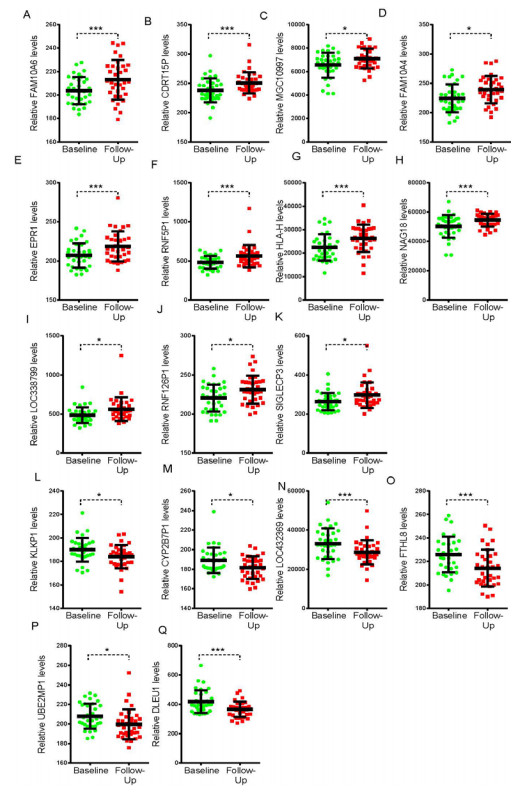
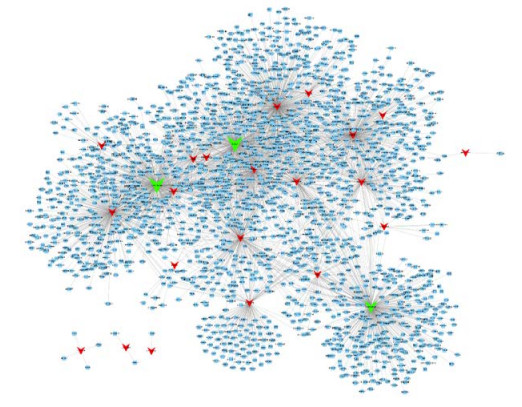
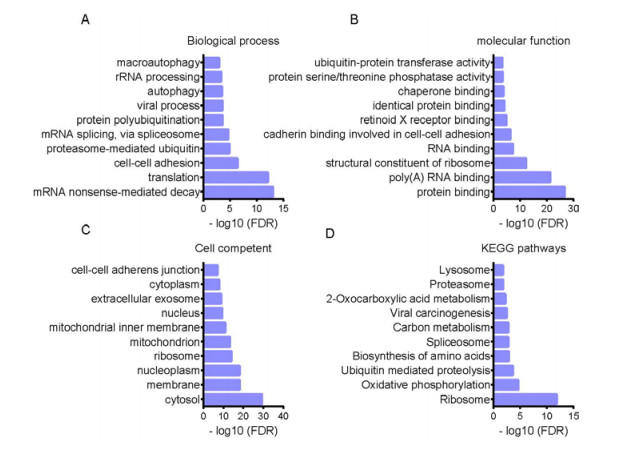
Figures(5)

Chunxiang Fan, Zouqin Huang, Binbin Chen, Baojin Chen, Qi Wang, Weidong Liu, Donghai Yu. Comprehensive analysis of key lncRNAs in ischemic stroke[J]. Mathematical Biosciences and Engineering, 2020, 17(2): 1318-1328. doi: 10.3934/mbe.2020066







 DownLoad:
DownLoad: