Citation: Lishui Shen, Xiaofeng Hu, Ting Chen, Guilin Shen, Dong Cheng. Integrated network analysis to explore the key mRNAs and lncRNAs in acute myocardial infarction[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 6426-6437. doi: 10.3934/mbe.2019321

| [1] | G. W. Reed, J. E. Rossi and C. P. Cannon, Acute myocardial infarction, The Lancet, 389 (2017), 197–210. |
| [2] | J. L. Anderson and D. A. Morrow, Acute myocardial infarction, N. Engl. J. Med., 376 (2017), 2053–2064. |
| [3] | C. Basso, S. Rizzo and G. Thiene, The metamorphosis of myocardial infarction following coronary recanalization, Cardiovasc. Pathol., 19 (2010), 22–28. |
| [4] | D. W. Kehl, N. Iqbal, A. Fard, et al., Biomarkers in acute myocardial injury, Transl. Res., 159 (2012), 252–264. |
| [5] | S. Korff, H. A. Katus and E. Giannitsis, Differential diagnosis of elevated troponins, Heart, 92 (2006), 987–993. |
| [6] | P. O'Brien, D. Smith, T. Knechtel, et al., Cardiac troponin I is a sensitive, specific biomarker of cardiac injury in laboratory animals, Lab. Anim-UK, 40 (2006), 153–171. |
| [7] | M. Saito, T. Ishimitsu, J. Minami, et al., Relations of plasma high-sensitivity C-reactive protein to traditional cardiovascular risk factors, Atherosclerosis, 167 (2003), 73–79. |
| [8] | V. A. Triant, H. Lee, C. Hadigan, et al., Increased acute myocardial infarction rates and cardiovascular risk factors among patients with human immunodeficiency virus disease, J. Clin. Endocr. Metab., 92 (2007), 2506–2512. |
| [9] | R. B. Ramos, C. M. Strunz, S. D. Avakian, et al., B-type natriuretic peptide as a predictor of anterior wall location in patients with non-ST-elevation myocardial infarction, Clinics, 66 (2011), 437–441. |
| [10] | O. M. Martins, V. F. Fonseca, I. Borges, et al., C-reactive protein predicts acute myocardial infarction in high-risk noncardiac and vascular surgery, Am. Heart Assoc., 2009. |
| [11] | P. D. Zamore and B. Haley, Ribo-gnome: The big world of small RNAs, Science, 309 (2005), 1519–1524. |
| [12] | T. Adachi, M. Nakanishi, Y. Otsuka, et al., Plasma microRNA 499 as a biomarker of acute myocardial infarction, Clin. Chem., 56 (2010), 1183–1185. |
| [13] | M. Vausort, D. R. Wagner and Y. Devaux, Long noncoding RNAs in patients with acute myocardial infarction novelty and significance, Circ. Res., 115 (2014), 668–677. |
| [14] | J. Liu, F. Sun, Y. Wang, et al., Suppression of microRNA-16 protects against acute myocardial infarction by reversing beta2-adrenergic receptor down-regulation in rats, Oncotarget, 8 (2017), 20122. |
| [15] | A. E. Kornienko, P. M. Guenzl, D. P. Barlow, et al., Gene regulation by the act of long non-coding RNA transcription, BMC biol., 11 (2013), 59. |
| [16] | K. C. Yang and J. M. Nerbonne, Mechanisms contributing to myocardial potassium channel diversity, regulation and remodeling, Trends Cardiovas. Med., 26 (2016), 209–218. |
| [17] | Y. Guo, F. Luo, Q. Liu, et al., Regulatory non-coding RNAs in acute myocardial infarction, J. Cell Mol. Med., (2016). |
| [18] | R. Suresh, X. Li, A. Chiriac, et al., Transcriptome from circulating cells suggests dysregulated pathways associated with long-term recurrent events following first-time myocardial infarction, J. Mol. Cell Cardiol., 74 (2014), 13–21. |
| [19] | X. Zhang, S. Sun, J. K. Pu, et al., Long non-coding RNA expression profiles predict clinical phenotypes in glioma, Neurobiol. Dis., 48 (2012), 1–8. |
| [20] | J. M. Wettenhalland G. K. Smyth, limmaGUI: A graphical user interface for linear modeling of microarray data, Bioinformatics, 20 (2004), 3705–3706. |
| [21] | D. W. Huang, B. T. Sherman and R. A. Lempicki, Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources, Nat. Protoc., 4 (2009), 44–57. |
| [22] | D. Zhang, W. Ma, Y. He, et al., Data of the interacting protein networks and nucleotide metabolism pathways related to NDK and NT5, Data Brief, 9 (2016), 1063–1066. |
| [23] | H. Yan, Z. Li, Q. Shen, et al., Aberrant expression of cell cycle and material metabolism-related genes contributes to hepatocellular carcinoma occurrence, Pathol. Res. Pract., 213 (2017), 316–321. |
| [24] | M. Kohl, S. Wiese and B. Warscheid, Cytoscape: Software for visualization and analysis of biological networks, Methods Mol. Biol., 696 (2011), 291–303. |
| [25] | D. Szklarczyk, A. Franceschini, S. Wyder, et al., STRING v10: protein-protein interaction networks, integrated over the tree of life, Nucleic. Acids Res., 43 (2014), D447–D452. |
| [26] | X. Wangand L. Liotta, Clinical bioinformatics: a new emerging science, J. clin. Bioinform., 1 (2011), 1. |
| [27] | J. R. Prensner and A. M. Chinnaiyan, The emergence of lncRNAs in cancer biology, Cancer Discov., 1 (2011), 391–407. |
| [28] | M. Huarte, The emerging role of lncRNAs in cancer, Nat. Med., 21 (2015), 1253. |
| [29] | A. Fatica and I. Bozzoni, Long non-coding RNAs: New players in cell differentiation and development, Nat. Rev. Genet., 15 (2014), 7. |
| [30] | Q. Shi and X. Yang, Circulating microRNA and long noncoding RNA as biomarkers of cardiovascular diseases, J. Cell Physiol., 231 (2016), 751–755. |
| [31] | K. Wang, C. Y. Liu, L. Y. Zhou, et al., APF lncRNA regulates autophagy and myocardial infarction by targeting miR-188-3p, Nat. Commun., 6 (2015), 6779. |
| [32] | Y. Zhang, L. Sun, L. Xuan, et al., Reciprocal changes of circulating long non-coding RNAs ZFAS1 and CDR1AS predict acute myocardial infarction, Sci. Rep-UK, 6 (2016). |
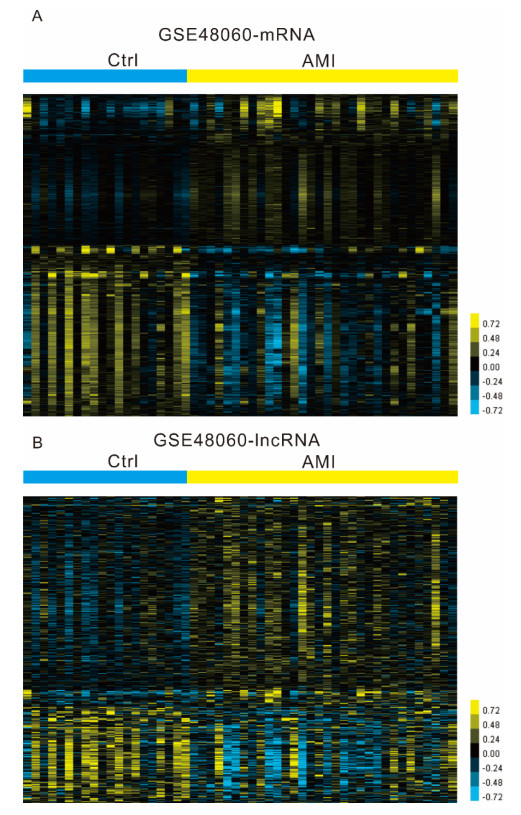
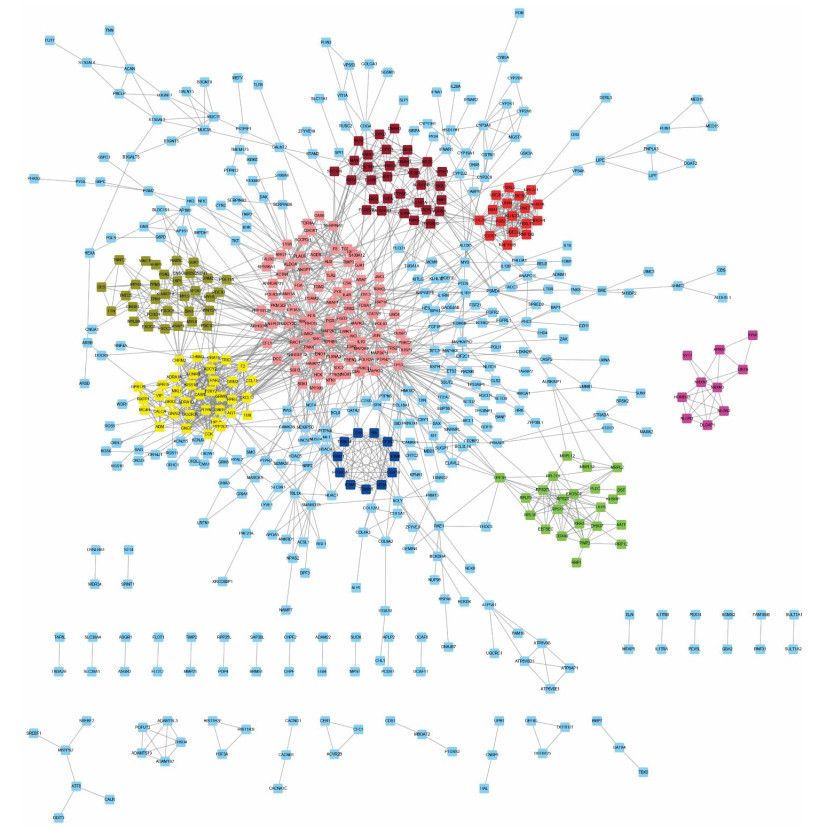

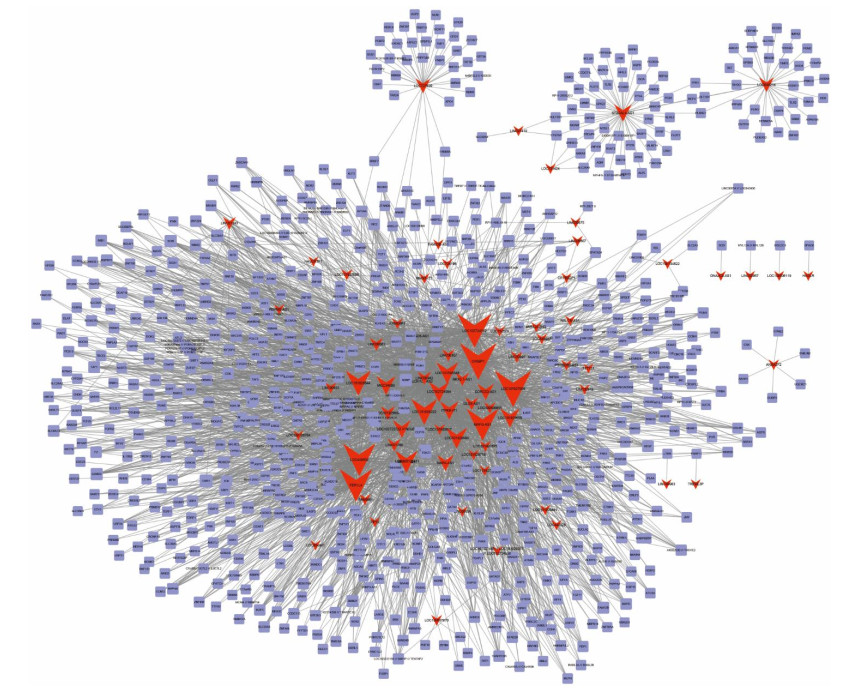
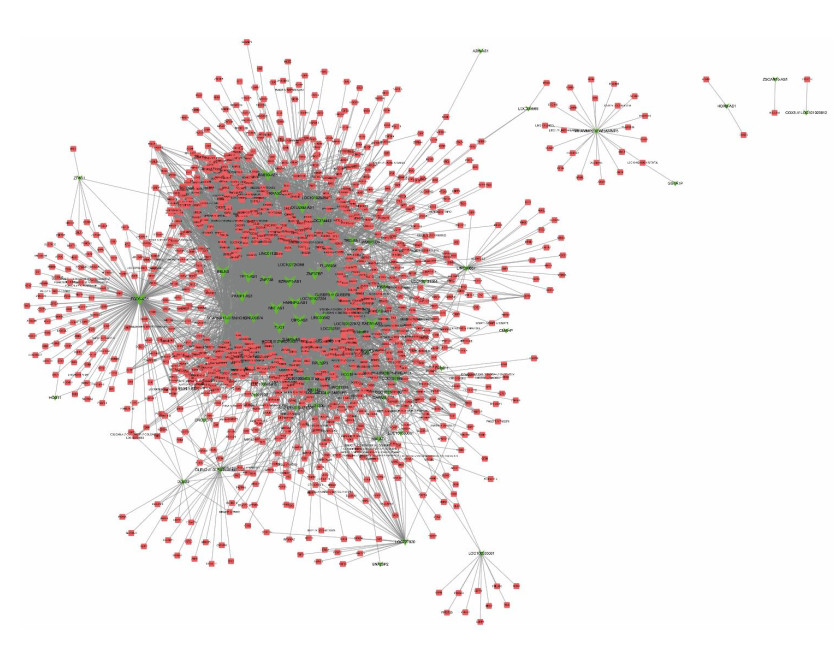
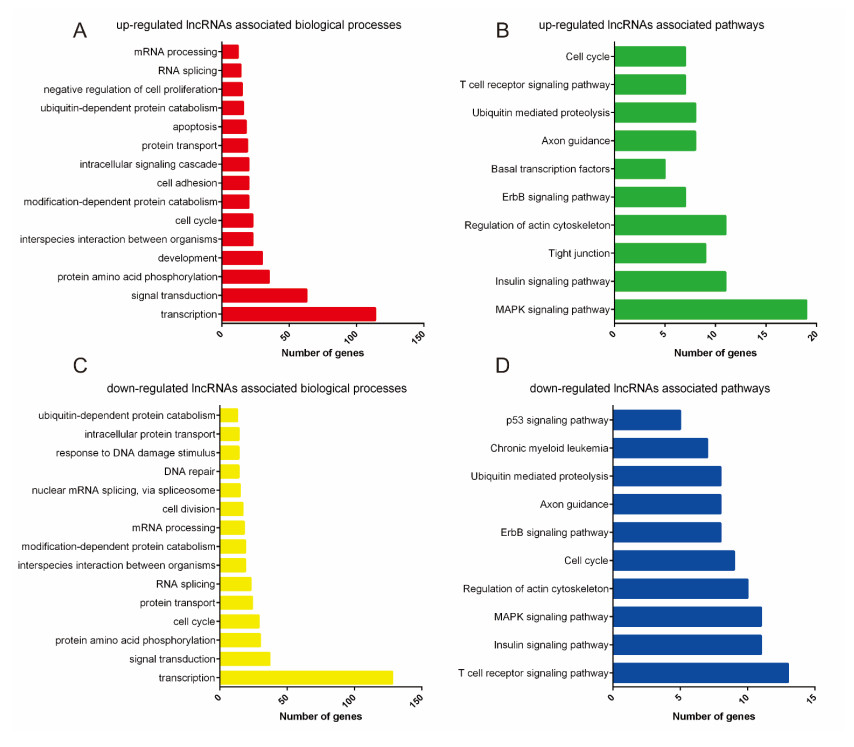
Figures(6)

Lishui Shen, Xiaofeng Hu, Ting Chen, Guilin Shen, Dong Cheng. Integrated network analysis to explore the key mRNAs and lncRNAs in acute myocardial infarction[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 6426-6437. doi: 10.3934/mbe.2019321







 DownLoad:
DownLoad: