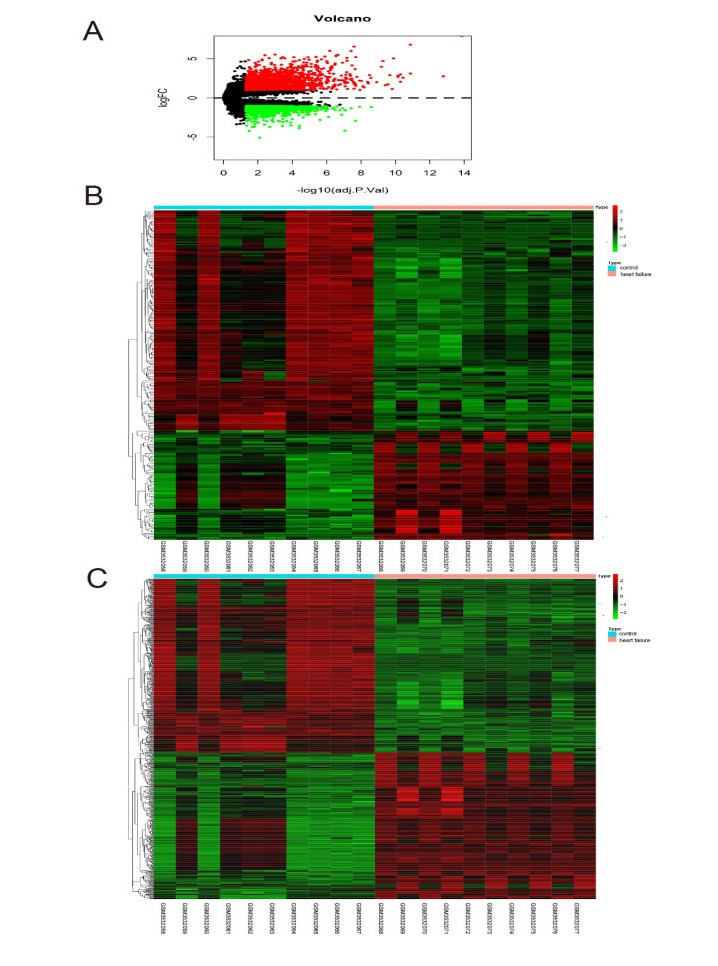
Heart failure (HF), a common disease in adults, accounts for significantly global morbidity and mortality. Due to population aging, therapeutic progression in acute cardiovascular events, the prevalence of HF is increasing, in spite of the efficacy of multiple therapies for HF patients with decreased ejection fraction. Despite great progress in the underlying molecular mechanisms, it remains incompletely clear of the function of competing endogenous RNA (ceRNA) network in HF pathogenesis. Herein, we established an HF-related ceRNA network on the basis of differentially expressed long noncoding RNAs (lncRNAs), microRNAs (miRNAs) as well as mRNAs from GSE136547 and GSE124401 datasets. In brief, the ceRNA network composed of 58 mRNA nodes, 5 miRNA nodes, 82 lncRNA nodes as well as 252 edges. In addition, three lncRNAs (KCNQ1OT1, XIST and AC010336) with higher node degrees than other lncRNAs were chosen as hub nodes. At the same time, we have established five subnetwork of miR-17-5p, miR-20b-5p, miR-107, miR-125a-5p and miR-140-5p centered ceRNA. Pathway analysis revealed the enrichment of ceRNA network in cell cycle pathways. Collectively, our research sheds new lights on the essential functions of ceRNA network in HF development, which also suggests possible application of these hub nodes as diagnostic biomarkers as well as therapeutic targets.
Citation: Xudan Ma, Qijun Zhang, Haihong Zhu, Kefeng Huang, Weina Pang, Qin Zhang. Establishment and analysis of the lncRNA-miRNA-mRNA network based on competitive endogenous RNA identifies functional genes in heart failure[J]. Mathematical Biosciences and Engineering, 2021, 18(4): 4011-4026. doi: 10.3934/mbe.2021201
Heart failure (HF), a common disease in adults, accounts for significantly global morbidity and mortality. Due to population aging, therapeutic progression in acute cardiovascular events, the prevalence of HF is increasing, in spite of the efficacy of multiple therapies for HF patients with decreased ejection fraction. Despite great progress in the underlying molecular mechanisms, it remains incompletely clear of the function of competing endogenous RNA (ceRNA) network in HF pathogenesis. Herein, we established an HF-related ceRNA network on the basis of differentially expressed long noncoding RNAs (lncRNAs), microRNAs (miRNAs) as well as mRNAs from GSE136547 and GSE124401 datasets. In brief, the ceRNA network composed of 58 mRNA nodes, 5 miRNA nodes, 82 lncRNA nodes as well as 252 edges. In addition, three lncRNAs (KCNQ1OT1, XIST and AC010336) with higher node degrees than other lncRNAs were chosen as hub nodes. At the same time, we have established five subnetwork of miR-17-5p, miR-20b-5p, miR-107, miR-125a-5p and miR-140-5p centered ceRNA. Pathway analysis revealed the enrichment of ceRNA network in cell cycle pathways. Collectively, our research sheds new lights on the essential functions of ceRNA network in HF development, which also suggests possible application of these hub nodes as diagnostic biomarkers as well as therapeutic targets.

| [1] | M. S. Islam, Heart failure: From research to clinical practice, Heart Failure: From Research to Clinical Practice, Springer 2018, pp. 1-3. |
| [2] |
S. L. James, D. Abate, K. H. Abate, S. M. Abay, C. Abbafati, N. Abbasi, et al., Global, regional, and national incidence, prevalence, and years lived with disability for 354 diseases and injuries for 195 countries and territories, 1990-2017: A systematic analysis for the Global Burden of Disease Study 2017, Lancet, 392 (2018), 1789-1858. doi: 10.1016/S0140-6736(18)32279-7

|
| [3] |
C. D. Kemp, J. V. Conte, The pathophysiology of heart failure, Cardiovasc. Pathol., 21 (2012), 365-371. doi: 10.1016/j.carpath.2011.11.007
|
| [4] |
S. M. Dunlay, S. A. Weston, S. J. Jacobsen, V. L. Roger, Risk factors for heart failure: A population-based case-control study, Am. J Med., 122 (2009), 1023-1028. doi: 10.1016/j.amjmed.2009.04.022
|
| [5] |
W. D. Rosamond, L. E. Chambless, G. Heiss, T. H. Mosley, J. Coresh, E. Whitsel, et al., Twenty-two-year trends in incidence of myocardial infarction, coronary heart disease mortality, and case fatality in 4 US communities, 1987-2008, Circulation, 125 (2012), 1848-1857. doi: 10.1161/CIRCULATIONAHA.111.047480
|
| [6] |
I. Baldi, D. Azzolina, P. Berchialla, D. Gregori, L. Scotti, G. Corrao, Comorbidity-adjusted relative survival in newly hospitalized heart failure patients: a population-based study, Int. J. Cardiol., 243 (2017), 385-388. doi: 10.1016/j.ijcard.2017.05.080
|
| [7] | A. E. Pasquinelli, MicroRNAs and their targets: Recognition, regulation and an emerging reciprocal relationship, Nat. Rev. Genet., 13 (2012), 271-282. |
| [8] | J. H. Noh, K. M. Kim, W. G. McClusky, K. Abdelmohsen, M. Gorospe, Cytoplasmic functions of long noncoding RNAs, Wiley Int. Rev. RNA, 9 (2018), e1471. |
| [9] |
F. Kopp, J. T. Mendell, Functional classification and experimental dissection of long noncoding RNAs, Cell, 172 (2018), 393-407. doi: 10.1016/j.cell.2018.01.011
|
| [10] | Y. Wang, X. Sun, The functions of LncRNA in the heart, Diabetes Res. Clin. Pract., 168 (2020). |
| [11] |
R. Kumarswamy, C. Bauters, I. Volkmann, F. Maury, J. Fetisch, A. Holzmann, et al., Circulating long noncoding RNA, LIPCAR, predicts survival in patients with heart failure, Circ. Res., 114 (2014), 1569-1575. doi: 10.1161/CIRCRESAHA.114.303915
|
| [12] |
S.-s. Zhou, J.-p. Jin, J.-q. Wang, Z.-g. Zhang, J.H. Freedman, Y. Zheng, et al., miRNAS in cardiovascular diseases: Potential biomarkers, therapeutic targets and challenges, Acta Pharmacol. Sin., 39 (2018), 1073-1084. doi: 10.1038/aps.2018.30
|
| [13] |
Y. Tay, J. Rinn, P. P. Pandolfi, The multilayered complexity of ceRNA crosstalk and competition, Nature, 505 (2014), 344-352. doi: 10.1038/nature12986
|
| [14] | T. Barrett, S. E. Wilhite, P. Ledoux, C. Evangelista, I. F. Kim, M. Tomashevsky, et al., NCBI GEO: Archive for functional genomics data sets—update, Nucleic Acids Res., 41 (2012), D991-D995. |
| [15] |
A. Jeggari, D. S. Marks, E. Larsson, miRcode: A map of putative microRNA target sites in the long non-coding transcriptome, Bioinformatics, 28 (2012), 2062-2063. doi: 10.1093/bioinformatics/bts344
|
| [16] | Y. Chen, X. Wang, miRDB: An online database for prediction of functional microRNA targets, Nucleic Acids Res., 48 (2020), D127-D131. |
| [17] | C.-H. Chou, S. Shrestha, C.-D. Yang, N.-W. Chang, Y.-L. Lin, K.-W. Liao, et al., miRTarBase update 2018: A resource for experimentally validated microRNA-target interactions, Nucleic Acids Res., 46 (2018), D296-D302. |
| [18] | V. Agarwal, G. W. Bell, J.-W. Nam, D. P. Bartel, Predicting effective microRNA target sites in mammalian mRNAs, eLife, 4 (2015), e05005. |
| [19] | D. Szklarczyk, A. L. Gable, D. Lyon, A. Junge, S. Wyder, J. Huerta-Cepas, et al., STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets, Nucleic Acids Res., 47 (2019), D607-D613. |
| [20] |
P. Ponikowski, S. D. Anker, K. F. AlHabib, M. R. Cowie, T. L. Force, S. Hu, et al., Heart failure: Preventing disease and death worldwide, ESC Heart Failure, 1 (2014), 4-25. doi: 10.1002/ehf2.12005
|
| [21] |
B. Ziaeian, G. C. Fonarow, Epidemiology and aetiology of heart failure, Nat Rev Cardiol., 13 (2016), 368-378. doi: 10.1038/nrcardio.2016.25
|
| [22] |
B. Tian, J. L. Manley, Alternative polyadenylation of mRNA precursors, Nat. Rev. Mol. Cell Biol., 18 (2017), 18-30. doi: 10.1038/nrm.2016.116
|
| [23] | J. Nourse, S. Spada, S. Danckwardt, Emerging roles of RNA 3'-end cleavage and polyadenylation in pathogenesis, diagnosis and therapy of human disorders, Biomolecules, 10 (2020), 915. |
| [24] | R. H. Deprez, A. F. Moorman, Expression of cell-cycle regulators during heart development and failure, Eur. Heart J., 20 (1999), 555-557. |
| [25] |
M. Hesse, A. Welz, B. K. Fleischmann, Heart regeneration and the cardiomyocyte cell cycle, Pflügers Archiv-Euro. J. Physiol., 470 (2018), 241-248. doi: 10.1007/s00424-017-2061-4
|
| [26] |
H. Ito, Cell cycle and heart failure, Nihon Yakurigaku Zasshi, 126 (2005), 381-384. doi: 10.1254/fpj.126.381
|
| [27] | C. Kanduri, Kcnq1ot1: A chromatin regulatory RNA, Elsevier, pp. 343-350. |
| [28] |
J. Rong, H. Pan, J. He, Y. Zhang, Y. Hu, C. Wang, et al., Long non-coding RNA KCNQ1OT1/microRNA-204-5p/LGALS3 axis regulates myocardial ischemia/reperfusion injury in mice, Cell. Signal., 66 (2020), 109441. doi: 10.1016/j.cellsig.2019.109441
|
| [29] |
Y. Jiang, W. Du, Q. Chu, Y. Qin, G. Tuguzbaeva, H. Wang, et al., Downregulation of long non-coding RNA Kcnq1ot1: an important mechanism of arsenic trioxide-induced long QT syndrome, Cell. Physiol. Biochem., 45 (2018), 192-202. doi: 10.1159/000486357
|
| [30] |
F. Yang, Y. Qin, J. Lv, Y. Wang, H. Che, X. Chen, et al., Silencing long non-coding RNA Kcnq1ot1 alleviates pyroptosis and fibrosis in diabetic cardiomyopathy, Cell Death Dis., 9 (2018), 1-13. doi: 10.1038/s41419-017-0012-9
|
| [31] | F. Yang, Y. Qin, Y. Wang, A. Li, J. Lv, X. Sun, et al., LncRNA KCNQ1OT1 mediates pyroptosis in diabetic cardiomyopathy, Cell Death Dis., 50 (2018), 1230-1244. |
| [32] |
L. Lai, Y. Xu, L. Kang, J. Yang, G. Zhu, LncRNA KCNQ1OT1 contributes to cardiomyocyte apoptosis by targeting FUS in heart failure, Exp. Mol. Pathol., 115 (2020), 104480. doi: 10.1016/j.yexmp.2020.104480
|
| [33] |
B. Liao, S. Dong, Z. Xu, F. Gao, S. Zhang, R. Liang, LncRNA Kcnq1ot1 renders cardiomyocytes apoptosis in acute myocardial infarction model by up-regulating Tead1, Life Sci., 256 (2020), 117811. doi: 10.1016/j.lfs.2020.117811
|
| [34] |
H. Liu, H. Deng, Y. Zhao, C. Li, Y. Liang, LncRNA XIST/miR-34a axis modulates the cell proliferation and tumor growth of thyroid cancer through MET-PI3K-AKT signaling, J. Exp. Clin. Cancer Res., 37 (2018), 1-12. doi: 10.1186/s13046-017-0664-4
|
| [35] |
J. Liu, L. Yao, M. Zhang, J. Jiang, M. Yang, Y. Wang, Downregulation of LncRNA-XIST inhibited development of non-small cell lung cancer by activating miR-335/SOD2/ROS signal pathway mediated pyroptotic cell death, Aging, 11 (2019), 7830. doi: 10.18632/aging.102291
|
| [36] | L. G. Yang, M. Z. Cao, J. Zhang, X. Y. Li, Q. L. Sun, LncRNA XIST modulates HIF‐1A/AXL signaling pathway by inhibiting miR‐93‐5p in colorectal cancer, Mol. Genet. Genom. Med., 8 (2020), e1112. |
| [37] |
L. Xiao, Y. Gu, Y. Sun, J. Chen, X. Wang, Y. Zhang, et al., The long noncoding RNA XIST regulates cardiac hypertrophy by targeting miR‐101, J. Cell. Physiol., 234 (2019), 13680-13692. doi: 10.1002/jcp.28047
|
| [38] | M. Zhang, H. Y. Liu, Y. L. Han, L. Wang, D. D. Zhai, T. Ma, et al., Silence of lncRNA XIST represses myocardial cell apoptosis in rats with acute myocardial infarction through regulating miR-449, Eur. Rev. Med. Pharmacol. Sci., 23 (2019), 8566-8572. |
| [39] |
Z. Li, Y. Zhang, N. Ding, Y. Zhao, Z. Ye, L. Shen, et al., Inhibition of lncRNA XIST improves myocardial I/R injury by targeting miR-133a through inhibition of autophagy and regulation of SOCS2, Mol. Therapy-Nucl., 18 (2019), 764-773. doi: 10.1016/j.omtn.2019.10.004
|
| [40] | D. P. Gómez-Mendoza, A. C. Lara-Ribeiro, T. Verano-Braga, Pathological cardiac remodeling seen by the eyes of proteomics, BBA, (2021), 140622. |
| [41] |
D. Wencker, M. Chandra, K. Nguyen, W. Miao, S. Garantziotis, S. M. Factor, et al., A mechanistic role for cardiac myocyte apoptosis in heart failure, J Clin. Invest., 111 (2003), 1497-1504. doi: 10.1172/JCI17664
|
| [42] |
A. Saraste, K. Pulkki, M. Kallajoki, P. Heikkilä , P. Laine, S. Mattila, et al., Cardiomyocyte apoptosis and progression of heart failure to transplantation, Eur. J. Clin. Invest., 29 (1999), 380-386. doi: 10.1046/j.1365-2362.1999.00481.x
|
| [43] |
P. M. Kang, S. Izumo, Apoptosis and heart failure: A critical review of the literature, Circ. Res., 86 (2000), 1107-1113. doi: 10.1161/01.RES.86.11.1107
|
| [44] |
G. Olivetti, R. Abbi, F. Quaini, J. Kajstura, W. Cheng, J. A. Nitahara, et al., Apoptosis in the failing human heart, N. Engl. J. Med., 336 (1997), 1131-1141. doi: 10.1056/NEJM199704173361603
|
| [45] |
J. Narula, N. Haider, R. Virmani, T. G. DiSalvo, F. D. Kolodgie, R. J. Hajjar, et al., Apoptosis in myocytes in end-stage heart failure, N. Engl. J. Med., 335 (1996), 1182-1189. doi: 10.1056/NEJM199610173351603
|
| [46] |
D. Zhang, V. Gaussin, G. E. Taffet, N. S. Belaguli, M. Yamada, R. J. Schwartz, et al., TAK1 is activated in the myocardium after pressure overload and is sufficient to provoke heart failure in transgenic mice, Nat. Med., 6 (2000), 556-563. doi: 10.1038/75037
|
| [47] |
J. Sadoshima, O. Montagne, Q. Wang, G. Yang, J. Warden, J. Liu, et al., The MEKK1-JNK pathway plays a protective role in pressure overload but does not mediate cardiac hypertrophy, J. Clin. Invest., 110 (2002), 271-279. doi: 10.1172/JCI0214938
|
| [48] | T. X. Lu, M. E. Rothenberg, MicroRNA, J. Allergy Clin. Immunol., 141 (2018), 1202-1207. |
| [49] |
J. Nourse, J. Braun, K. Lackner, S. Hüttelmaier, S. Danckwardt, Large‐scale identification of functional micro RNA targeting reveals cooperative regulation of the hemostatic system, J. Thromb. Haemost., 16 (2018), 2233-2245. doi: 10.1111/jth.14290
|
| [50] | A. Wojciechowska, A. Braniewska, K. Kozar-Kamińska, MicroRNA in cardiovascular biology and disease, Adv. Clin. Exp. Med., 26 (2017), 865-874. |
| [51] | D. A. Chistiakov, A. N. Orekhov, Y. V. Bobryshev, Cardiac-specific miRNA in cardiogenesis, heart function, and cardiac pathology (with focus on myocardial infarction), J. Mol. Cell. Cardiol., 94 (2016), 107-121. |
| [52] |
E. L. Vegter, P. van der Meer, L. J. de Windt, Y. M. Pinto, A. A. Voors, MicroRNAs in heart failure: From biomarker to target for therapy, Eur. J. Heart Fail., 18 (2016), 457-468. doi: 10.1002/ejhf.495
|
| [53] |
Z. Zhou, S. Chen, Z. Tian, S. Deng, X. Yi, S. Yang, et al., miR-20b-5p attenuates hypoxia-induced apoptosis in cardiomyocytes via the HIF-1 α/NF-κ B pathway, Acta Bioch. Biophys. Sinica, 52 (2020), 927-934. doi: 10.1093/abbs/gmaa056
|
| [54] | F. Tian, J. Wang, Z. Zhang, J. Yang, miR-107 modulates chondrocyte proliferation, apoptosis, and extracellular matrix synthesis by targeting PTEN, Int. J. Clin. Exp. Pathol., 12 (2019), 488. |
| [55] |
D. Lu, T. Thum, RNA-based diagnostic and therapeutic strategies for cardiovascular disease, Nat. Rev. Cardiol., 16 (2019), 661-674. doi: 10.1038/s41569-019-0218-x
|
| [56] | J. Nourse, S. Danckwardt, A novel rationale for targeting FXI: Insights from the hemostatic miRNA targetome for emerging anticoagulant strategies, Pharmacol. Ther., (2020), 107676. |





Figures(6) / Tables(4)

Xudan Ma, Qijun Zhang, Haihong Zhu, Kefeng Huang, Weina Pang, Qin Zhang. Establishment and analysis of the lncRNA-miRNA-mRNA network based on competitive endogenous RNA identifies functional genes in heart failure[J]. Mathematical Biosciences and Engineering, 2021, 18(4): 4011-4026. doi: 10.3934/mbe.2021201







 DownLoad:
DownLoad: