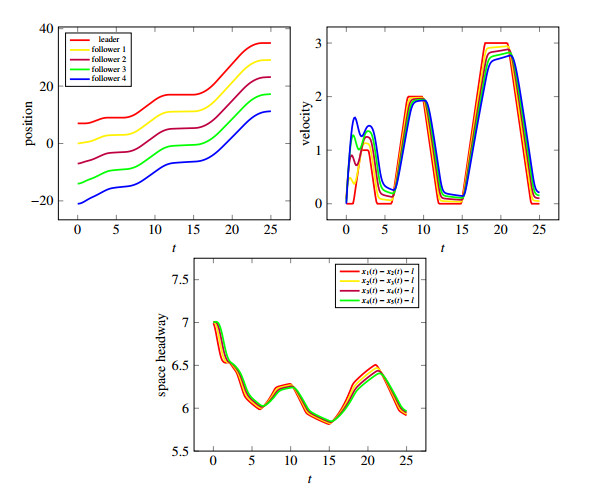
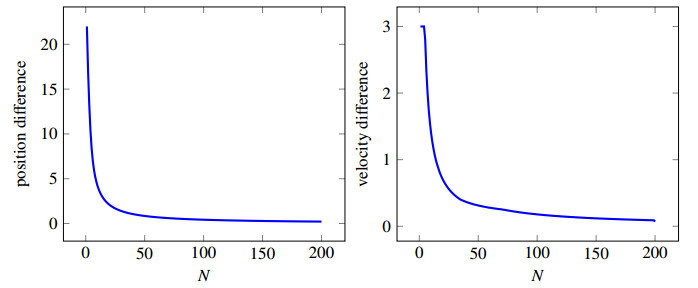
In this contribution we study the "Bando-follow the leader" car-following model, a second order ordinary differential equation, for its well-posedness. Under suitable conditions, we provide existence and uniqueness results, and also bounds on the higher derivatives, i.e., velocity and acceleration. We then extend the result to the "reaction" delay case where the delay is instantiated in reacting on the leading vehicle's position and velocity. We prove that the solution of the delayed model converges to the undelayed when the delay converges to zero and present some numerical examples underlying the idea that it is worth looking in more details into delay as it might explain problems in traffic flow like "phantom shocks" and "stop and go" waves.
Citation: Xiaoqian Gong, Alexander Keimer. On the well-posedness of the "Bando-follow the leader" car following model and a time-delayed version[J]. Networks and Heterogeneous Media, 2023, 18(2): 775-798. doi: 10.3934/nhm.2023033
In this contribution we study the "Bando-follow the leader" car-following model, a second order ordinary differential equation, for its well-posedness. Under suitable conditions, we provide existence and uniqueness results, and also bounds on the higher derivatives, i.e., velocity and acceleration. We then extend the result to the "reaction" delay case where the delay is instantiated in reacting on the leading vehicle's position and velocity. We prove that the solution of the delayed model converges to the undelayed when the delay converges to zero and present some numerical examples underlying the idea that it is worth looking in more details into delay as it might explain problems in traffic flow like "phantom shocks" and "stop and go" waves.

| [1] |
S. Albeaik, A. Bayen, M. T. Chiri, X. Gong, A. Hayat, N. Kardous, et al., Limitations and improvements of the intelligent driver model (IDM), SIAM J. Appl. Dyn. Syst., 21 (2022), 1862–1892. https://doi.org/10.1137/21M1406477 doi: 10.1137/21M1406477

|
| [2] |
I. Bae, J. Moon and J. Seo, Toward a comfortable driving experience for a self-driving shuttle bus, Electronics, 8 (2019), 943. https://doi.org/10.3390/electronics8090943 doi: 10.3390/electronics8090943
|
| [3] |
M. Bando, K. Hasebe, K. Nakanishi, A. Nakayama, A. Shibata, Y. Sugiyama, Phenomenological study of dynamical model of traffic flow, J. Phys. I, 5 (1995), 1389–1399. https://doi.org/10.1051/jp1:1995206 doi: 10.1051/jp1:1995206
|
| [4] |
M. Bando, K. Hasebe, A. Nakayama, A. Shibata, Y. Sugiyama, Dynamical model of traffic congestion and numerical simulation, Phys. Rev. E, 51 (1995), 1035–1042. https://doi.org/10.1103/PhysRevE.51.1035 doi: 10.1103/PhysRevE.51.1035
|
| [5] |
M. Bando, K. Hasebe, K. Nakanishi, A. Nakayama, Analysis of optimal velocity model with explicit delay, Phys. Rev. E, 58 (1998), 5429–5435. https://doi.org/10.1103/PhysRevE.58.5429 doi: 10.1103/PhysRevE.58.5429
|
| [6] |
S. Bexelius, An extended model for car-following, Transp. Res., 2 (1968), 13–21. https://doi.org/10.1016/0041-1647(68)90004-X doi: 10.1016/0041-1647(68)90004-X
|
| [7] |
P. Bosetti, M. Da Lio, A. Saroldi, On the human control of vehicles: an experimental study of acceleration, Eur. Transport Res. Rev., 6 (2014), 157–170. https://doi.org/10.1007/s12544-013-0120-2 doi: 10.1007/s12544-013-0120-2
|
| [8] |
M. Brackstone, M. McDonald, Car-following: A historical review, Transp. Res. Part F Psychol. Behav., 2 (1999), 181–196. https://doi.org/10.1016/S1369-8478(00)00005-X doi: 10.1016/S1369-8478(00)00005-X
|
| [9] | H. Brezis, Functional Analysis, Sobolev Spaces and Partial Differential Equations, New York: Springer, 2011. https://doi.org/10.1007/978-0-387-70914-7 |
| [10] |
M. Burger, S. Göttlich, T. Jung, Derivation of a first order traffic flow model of lighthill-whitham-richards type, IFAC-PapersOnLine, 51 (2018), 49–54. https://doi.org/10.1016/j.ifacol.2018.07.009 doi: 10.1016/j.ifacol.2018.07.009
|
| [11] |
R. E. Chandler, R. Herman, E. W. Montroll, Traffic dynamics: Studies in car following, Oper. Res., 6 (1958), 165–184. https://doi.org/10.1287/opre.6.2.165 doi: 10.1287/opre.6.2.165
|
| [12] |
L. Davis, Modifications of the optimal velocity traffic model to include delay due to driver reaction time, Physica A, 319 (2003), 557–567. https://doi.org/10.1016/S0378-4371(02)01457-7 doi: 10.1016/S0378-4371(02)01457-7
|
| [13] | R. D. Driver, Ordinary and delay differential equations, Springer Science Business Media, 2012. |
| [14] | J. Eggert, F. Damerow, S. Klingelschmitt, The foresighted driver model, 2015 IEEE Intelligent Vehicles Symposium (IV), IEEE, Seoul, Korea, (2015), 322–329. https://doi.org/10.1109/IVS.2015.7225706 |
| [15] |
D. C. Gazis, R. Herman, R. W. Rothery, Nonlinear follow-the-leader models of traffic flow, Oper. Res., 9 (1961), 545–567. https://doi.org/10.1287/opre.9.4.545 doi: 10.1287/opre.9.4.545
|
| [16] |
D. Gazis, R. Herman, The moving and "phantom" bottlenecks, Transp. Sci., 26 (1992), 223–229. https://doi.org/10.1287/trsc.26.3.223 doi: 10.1287/trsc.26.3.223
|
| [17] | X. Gong, B. Piccoli, G. Visconti, Mean-field limit of a hybrid system for multi-lane multi-class traffic, 2020. https://doi.org/10.48550/arXiv.2007.14655 |
| [18] | P. Hartman, Ordinary Differential Equations: Second Edition, Classics in Applied Mathematics, Society for Industrial and Applied Mathematics (SIAM, 3600 Market Street, Floor 6, Philadelphia, PA 19104), 1982. |
| [19] |
S. Hoogendoorn, P. Bovy, State-of-the-art of vehicular traffic flow modelling, Proc. Inst. Mech. Eng., Part I: J. Syst. Control Eng., 215 (2001), 283–303. https://doi.org/10.1177/095965180121500402 doi: 10.1177/095965180121500402
|
| [20] |
R. Jiang, M. Hu, B. Jia, R. Wang, Q. Wu, The effects of reaction delay in the nagel-schreckenberg traffic flow model, Eur. Phys. J. B, 54 (2006), 267–273. https://doi.org/10.1140/epjb/e2006-00449-5 doi: 10.1140/epjb/e2006-00449-5
|
| [21] | N. Kardous, A. Hayat, S. McQuade, X. Gong, S. Truong, P. Arnold, et al., A rigorous multi-population multi-lane hybrid traffic model and its mean-field limit for dissipation of waves via autonomous vehicles, 2022. https://doi.org/10.48550/arXiv.2205.06913 |
| [22] | R. Keane, H. O. Gao, Calibration for traffic microsimulation using trajectory data, part 1: Fast calibration using the adjoint method, CoRR, 2019. https://doi.org/10.48550/arXiv.1901.06452 |
| [23] |
A. Keimer, L. Pflug, Nonlocal conservation laws with time delay, Nonlinear Differ. Equations Appl., 26 (2019), 1–34. https://doi.org/10.1007/s00030-019-0597-z doi: 10.1007/s00030-019-0597-z
|
| [24] |
A. Keimer, L. Pflug, M. Spinola, Existence, uniqueness and regularity of multi-dimensional nonlocal balance laws with damping, J. Math. Anal. Appl., 466 (2018), 18–55. https://doi.org/10.1016/j.jmaa.2018.05.013 doi: 10.1016/j.jmaa.2018.05.013
|
| [25] |
A. Kesting, M. Treiber, D. Helbing, Enhanced intelligent driver model to access the impact of driving strategies on traffic capacity, Phil. Trans. R. Soc. A, 368 (2010), 4585–4605. https://doi.org/10.1098/rsta.2010.0084 doi: 10.1098/rsta.2010.0084
|
| [26] |
Z. Li, W. Li, S. Xu, Y. Qian, Stability analysis of an extended intelligent driver model and its simulations under open boundary condition, Physica A, 419 (2015), 526–536. https://doi.org/10.1016/j.physa.2014.10.063 doi: 10.1016/j.physa.2014.10.063
|
| [27] | M. Liebner, M. Baumann, F. Klanner, C. Stiller, Driver intent inference at urban intersections using the intelligent driver model, in 2012 IEEE Intelligent Vehicles Symposium, IEEE, Madrid, Spain, (2012), 1162–1167. https://doi.org/10.1109/IVS.2012.6232131 |
| [28] |
B. N. Matcha, S. Namasivayam, M. H. Fouladi, K. C. Ng, S. Sivanesan, S. Y. E. Noum, Simulation strategies for mixed traffic conditions: A review of car-following models and simulation frameworks, J. Eng., 2020 (2020), 1–22. https://doi.org/10.1155/2020/8231930 doi: 10.1155/2020/8231930
|
| [29] |
K. Nagel, M. Schreckenberg, A cellular automaton model for freeway traffic, J. Phys. I, 2 (1992), 2221–2229. https://doi.org/10.1051/jp1:1992277 doi: 10.1051/jp1:1992277
|
| [30] |
G. Orosz, B. Krauskopf, R. E. Wilson, Bifurcations and multiple traffic jams in a car-following model with reaction-time delay, Physica D, 211 (2005), 277–293. https://doi.org/10.1016/j.physd.2005.09.004 doi: 10.1016/j.physd.2005.09.004
|
| [31] |
G. Orosz, G. Stépán, Subcritical hopf bifurcations in a car-following model with reaction-time delay, Proc. Math. Phys. Eng. Sci., 462 (2006), 2643–2670. https://doi.org/10.1098/rspa.2006.1660 doi: 10.1098/rspa.2006.1660
|
| [32] |
Oussama, T. Peter, H. Zebiri, B. Mourllion, M. Basset, Modified intelligent driver model for driver safety and traffic stability improvement, IFAC Proceedings Volumes, 46 (2013), 744–749. https://doi.org/10.3182/20130904-4-JP-2042.00132 doi: 10.3182/20130904-4-JP-2042.00132
|
| [33] |
B. Seibold, M. R. Flynn, A. R. Kasimov, R. R. Rosales, Constructing set-valued fundamental diagrams from jamiton solutions in second order traffic models, Networks Heterogen. Media, 8 (2013), 745–772. https://doi.org/10.3934/nhm.2013.8.745 doi: 10.3934/nhm.2013.8.745
|
| [34] |
R. E. Stern, S. Cui, M. L. Delle Monache, R. Bhadani, M. Bunting, M. Churchill, et al. Dissipation of stop-and-go waves via control of autonomous vehicles: Field experiments, Transp. Res. Part C Emerging Technol., 89 (2018), 205–221. https://doi.org/10.1016/j.trc.2018.02.005 doi: 10.1016/j.trc.2018.02.005
|
| [35] |
M. Treiber, A. Hennecke, D. Helbing, Congested traffic states in empirical observations and microscopic simulations, Phys. Rev. E, 62 (2000), 1805. https://doi.org/10.1103/PhysRevE.62.1805 doi: 10.1103/PhysRevE.62.1805
|
| [36] | M. Treiber, A. Kesting, Traffic flow dynamics: Data, models and simulation, Berlin Heidelberg: Springer-Verlag, 2013. |
| [37] |
L. Yu, Z. Ke Shi, T. Li, A new car-following model with two delays, Phys. Lett. A, 378 (2014), 348–357. https://doi.org/10.1016/j.physleta.2013.11.030 doi: 10.1016/j.physleta.2013.11.030
|
| [38] | C. Zhai, W. Liu, L. Huang, F. Tan, Feedback control strategy in a car-following model with two delays, in 2016 12th World Congress on Intelligent Control and Automation (WCICA), IEEE, Guilin, (2016), 2951–2956. https://doi.org/10.1109/WCICA.2016.7578259 |
| [39] |
H. Zhu, S. Dai, Analysis of car-following model considering driver's physical delay in sensing headway, Physica A, 387 (2008), 3290–3298. https://doi.org/10.1016/j.physa.2008.01.103 doi: 10.1016/j.physa.2008.01.103
|
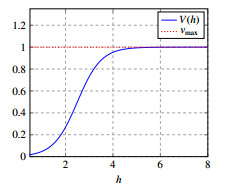
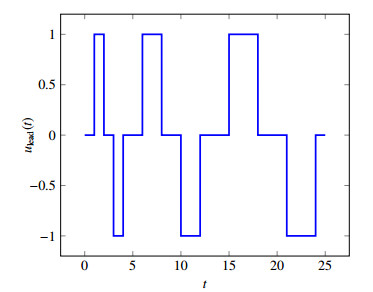
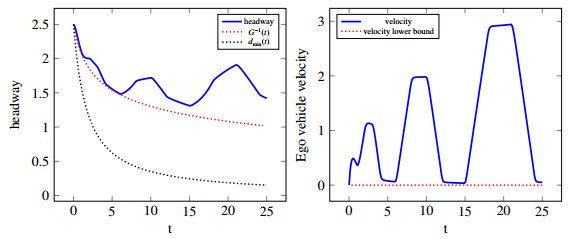
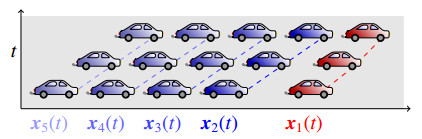
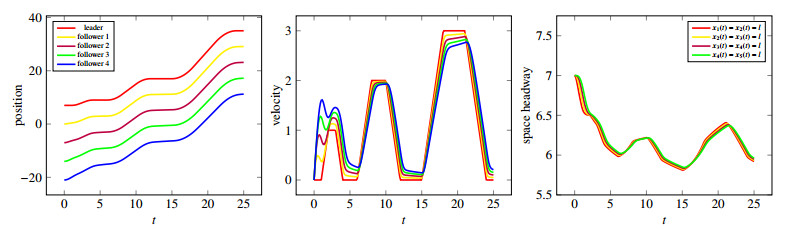
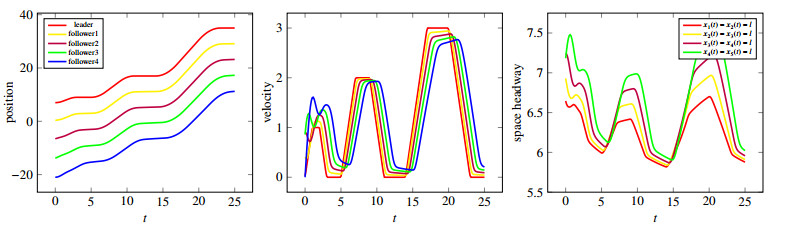
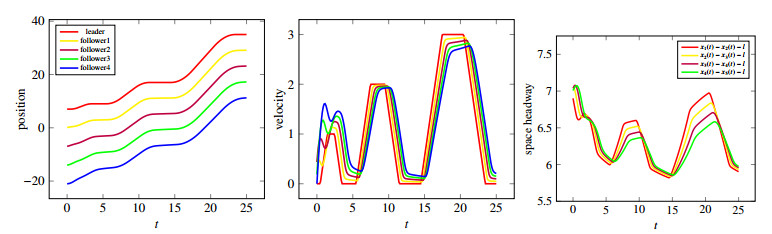
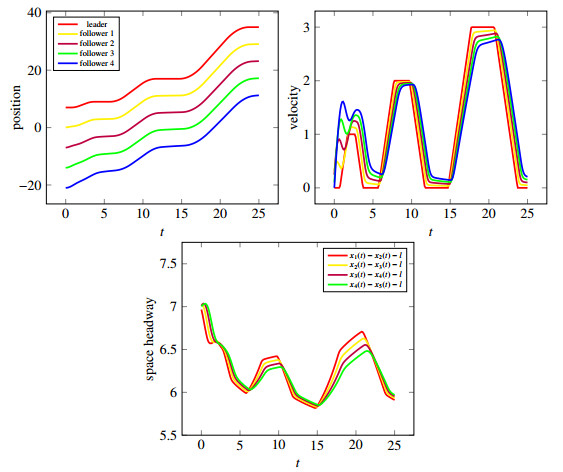
Figures(10)
Xiaoqian Gong, Alexander Keimer. On the well-posedness of the "Bando-follow the leader" car following model and a time-delayed version[J]. Networks and Heterogeneous Media, 2023, 18(2): 775-798. doi: 10.3934/nhm.2023033







 DownLoad:
DownLoad: