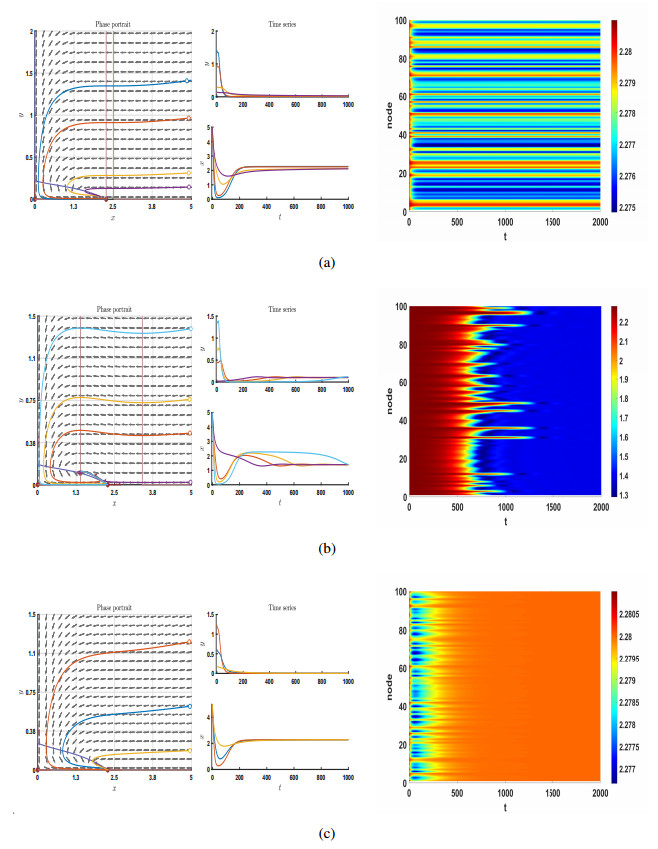
This paper used a Holling-IV nutrient-plankton model with a network to describe algae's spatial and temporal distribution and variation in a specific sea area. The stability and bifurcation of the nonlinear dynamic model of harmful algal blooms (HABs) were analyzed using the nonlinear dynamic theory and de-eutrophication's effect on algae's nonlinear dynamic behavior. The conditions for equilibrium points (local and global), saddle-node, transcritical, Hopf-Andronov and Bogdanov-Takens (B-T) bifurcation were obtained. The stability of the limit cycle was then judged and the rich and complex phenomenon was obtained by numerical simulations, which revealed the robustness of the nutrient-plankton system by switching between nodes. Also, these results show the relationship between HABs and bifurcation, which has important guiding significance for solving the environmental problems of HABs caused by the abnormal increase of phytoplankton.
Citation: Wenjie Yang, Qianqian Zheng, Jianwei Shen, Linan Guan. Bifurcation and pattern dynamics in the nutrient-plankton network[J]. Mathematical Biosciences and Engineering, 2023, 20(12): 21337-21358. doi: 10.3934/mbe.2023944
This paper used a Holling-IV nutrient-plankton model with a network to describe algae's spatial and temporal distribution and variation in a specific sea area. The stability and bifurcation of the nonlinear dynamic model of harmful algal blooms (HABs) were analyzed using the nonlinear dynamic theory and de-eutrophication's effect on algae's nonlinear dynamic behavior. The conditions for equilibrium points (local and global), saddle-node, transcritical, Hopf-Andronov and Bogdanov-Takens (B-T) bifurcation were obtained. The stability of the limit cycle was then judged and the rich and complex phenomenon was obtained by numerical simulations, which revealed the robustness of the nutrient-plankton system by switching between nodes. Also, these results show the relationship between HABs and bifurcation, which has important guiding significance for solving the environmental problems of HABs caused by the abnormal increase of phytoplankton.

| [1] |
Q. Zheng, J. Shen, V. Pandey, Y. Zhao, L. Guan, Spatiotemporal dynamics of periodic waves in SIR model with driving factors, New J. Phys., 25 (2023), 063028. https://doi.org/10.1088/1367-2630/acdb91 doi: 10.1088/1367-2630/acdb91

|
| [2] |
L. Pei, M. Zhang, Long-term predictions of current confirmed and dead cases of COVID-19 in China by the non-autonomous delayed epidemic models, Cognit. Neurodyn., 16 (2022), 229–238. https://doi.org/10.1007/s11571-021-09701-1 doi: 10.1007/s11571-021-09701-1
|
| [3] |
Q. Zheng, J. Shen, V. Pandey, L. Guan, Y. Guo, Turing instability in a network-organized epidemic model with delay, Chaos Solitons Fractals, 168 (2023), 113205. https://doi.org/10.1016/j.chaos.2023.113205 doi: 10.1016/j.chaos.2023.113205
|
| [4] |
Q. Zheng, J. Shen, L. Zhou, L Guan, Turing pattern induced by the directed ER network and delay, Math. Biosci. Eng., 19 (2022), 11854–11867. https://doi.org/10.3934/mbe.2022553 doi: 10.3934/mbe.2022553
|
| [5] |
N. Melek Manshouri, Identifying COVID-19 by using spectral analysis of cough recordings: a distinctive classification study, Cognit. Neurodyn., 16 (2022), 239–253. https://doi.org/10.1007/s11571-021-09695-w doi: 10.1007/s11571-021-09695-w
|
| [6] |
Q. Zheng, V. Pandey, J. Shen, Y. Xu, L. Guan, Pattern dynamics in the epidemic model with diffusion network, Europhys. Lett., 137 (2022), 42002. https://doi.org/10.1209/0295-5075/ac58bd doi: 10.1209/0295-5075/ac58bd
|
| [7] |
Q. Zheng, J. Shen, Y. Xu, V. Pandey, L. Guan, Pattern mechanism in stochastic SIR networks with ER connectivity, Phys. A Stat. Mech. Appl., 603 (2022), 127765. https://doi.org/10.1016/j.physa.2022.127765 doi: 10.1016/j.physa.2022.127765
|
| [8] |
Z. Song, J. Xu, Stability switches and double Hopf bifurcation in a two-neural network system with multiple delays, Cognit. Neurodyn., 7 (2013), 505–521. https://doi.org/10.1007/s11571-013-9254-0 doi: 10.1007/s11571-013-9254-0
|
| [9] |
R. S. Kumar, G. Sugumaran, R. Raja, Q. Zhu, U. K. Raja, New stability criterion of neural networks with leakage delays and impulses: a piecewise delay method, Cognit. Neurodyn., 10 (2016), 85–98. https://doi.org/10.1007/s11571-015-9356-y doi: 10.1007/s11571-015-9356-y
|
| [10] |
Y. Yang, F. Meng, Y. Xu, Global bifurcation analysis in a predator-prey system with simplified Holling IV functional response and antipredator behavior, Math. Methods Appl. Sci., 46 (2023), 6135–6153. https://doi.org/10.1002/mma.8896 doi: 10.1002/mma.8896
|
| [11] |
Z. Shang, Y. Qiao, Multiple bifurcations in a predator-prey system of modified Holling and Leslie type with double Allee effect and nonlinear harvesting, Math. Comput. Simul., 205 (2023), 745–764. https://doi.org/10.1016/j.matcom.2022.10.028 doi: 10.1016/j.matcom.2022.10.028
|
| [12] | D. Mukherjee, Fear induced dynamics on Leslie-Gower predator-prey system with Holling-type IV functional response, Jambura J. Biomath., 3 (2022). https://doi.org/10.34312/jjbm.v3i2.14348 |
| [13] |
S. Sarwardi, S. Hossain, F. Al Basir, S. Ray, Mathematical analysis of an ecological system using a non-monotonic functional response: effects of gestation delay and predator harvesting, Int. J. Dyn. Control, 8 (2022), 1–14. https://doi.org/10.1007/s40435-022-00999-1 doi: 10.1007/s40435-022-00999-1
|
| [14] |
G. Ble, V. Castellanos, M. A. Dela-Rosa, Coexistence of species in a tritrophic food chain model with Holling functional response type IV, Math. Methods Appl. Sci., 41 (2018), 6683–6701. https://doi.org/10.1002/mma.5184 doi: 10.1002/mma.5184
|
| [15] |
J. Song, Y. Xia, Y. Bai, Y. Cai, D O'Regan, A non-autonomous Leslie-Gower model with Holling type IV functional response and harvesting complexity, Adv. Differ. Equations, 2019 (2019), 1–12. https://doi.org/10.1186/s13662-019-2203-4 doi: 10.1186/s13662-019-2203-4
|
| [16] |
S. Karthikeyan, P. Ramesh, M. Sambath, Stability analysis of harvested fractional-order prey-predator model with Holling type IV response, Int. J. Nonlinear Anal. Appl., 14 (2023), 2019–2030. https://doi.org/10.22075/IJNAA.2022.27700.3685 doi: 10.22075/IJNAA.2022.27700.3685
|
| [17] | R. N. Shalan, Global stability of three species food web with Holling type IV functional response, J. Coll. Edu., 5 (2015), 315–330. https://www.iasj.net/iasj/article/128178 |
| [18] | D. B. Prakash, D. K. K. Vamsi, Stochastic time-optimal control studies for additional food provided prey-predator systems involving Holling type-IV functional response, preprint, arXiv: 2212.06447. |
| [19] |
Shivam, M. Kumar, T. Singh, R. Dubey, K. Singh, Analytical study of food-web system via turing patterns, AIP Confer. Proc., 2481 (2022), 040036. https://doi.org/10.1063/5.0103837 doi: 10.1063/5.0103837
|
| [20] | R. C. Yu, S. H. Lü, Y. B. Liang, Harmful algal blooms in the coastal waters of China, Global Ecol. Oceanogr. Harmful Algal Blooms Ecol. Stud., 232 (2018), 309–316. https://doi.org/10.1007/978-3-319-70069-4 15 |
| [21] | Z. Yu, X. Song, X. Cao, Y. Liu, Mitigation and control of harmful algal blooms, Global Ecol. Oceanogr. Harmful Algal Blooms Ecol. Stud., 232 (2018), 403–423. https://doi.org/10.1007/978-3-319-70069-4 21 |
| [22] | H. L. Wang, J. F. Feng, HABs Ecosystem Dynamics and Prediction, Tianjin University Press, Tianjin, 2006. https://doi.org/7-5618-2275-8 |
| [23] | Q. C. Zhang, H. L. Wang, Z. Lan, F. Shen, A. Ren, H. Liu, Bifurcation and Chaos Theory and Applications, Tianjin University Press, Tianjin, 2005. |
| [24] |
J. C. Huang, D. M. Xiao, Analyses of bifurcations and stability in a predator-prey system with Holling type IV functional response, Acta Math. Appl. Sin., 20 (2004), 167–178. https://doi.org/10.1007/s10255-004-0159-x doi: 10.1007/s10255-004-0159-x
|
| [25] | U. Sommer, Plankton Ecology: Succession in PlanktonCommunities, Springer, Berlin, 1989. |
| [26] |
K. G. Sellner, Physiology ecology and toxic properties of marine cyanobacteria blooms, Limnol. Oceanogr., 42 (1997), 1089–1104. https://doi.org/10.4319/lo.1997.42.5part2.1089 doi: 10.4319/lo.1997.42.5part2.1089
|
| [27] |
D. M. Anderson, Bloom dynamics of toxic Alexandrium species in the northeastem US, Limnol. Oceanogr., 42 (1997), 1009–1022. https://doi.org/10.4319/lo.1997.42.5part2.1009 doi: 10.4319/lo.1997.42.5part2.1009
|
| [28] |
A. Tester, K. A. Steidinger, Gymnodinium breve red tide blooms: initiation transport and conse-quences of surface circulation, Limnol. Oceanogr., 42 (1997), 1039–1051. https://doi.org/10.4319/lo.1997.42.5_part_2.1039 doi: 10.4319/lo.1997.42.5_part_2.1039
|
| [29] |
W. J. Yang, Q. Q. Zheng, J. W. Shen, Pattern dynamics and bifurcation in delayed SIR network with diffusion network, Int. J. Biomath., 2023 (2023), 1–18. https://doi/10.1142/S1793524523500146 doi: 10.1142/S1793524523500146
|
| [30] |
C. Tian, S. G. Ruan, Pattern formation and synchronism in an allelopathic plankton model with delay in a network, SIAM J. Appl. Dyna. Syst., 18 (2019), 531–557. https://doi.org/10.1137/18M1204966 doi: 10.1137/18M1204966
|
| [31] | Y. Kuznetsov, Elements of Applied Bifurcation Theory,, 2$^{st}$ edition, Springer-Verlag, New York, 1998. |
| [32] | D. M. Xiao, S. G. Ruan, Bogdanov-Takens bifurcations in predator-prey systems with constant rate harvesting, Fields Inst. Commun., 21 (1999), 493–506. |







Figures(8)

Wenjie Yang, Qianqian Zheng, Jianwei Shen, Linan Guan. Bifurcation and pattern dynamics in the nutrient-plankton network[J]. Mathematical Biosciences and Engineering, 2023, 20(12): 21337-21358. doi: 10.3934/mbe.2023944







 DownLoad:
DownLoad: