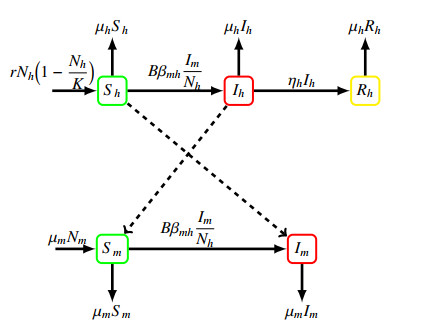
Dengue is a deadly illness in almost all parts of Indonesia, including East Java. This paper analyzes the dengue transmission model by considering the recruitment rate in the form of the logistic growth of the human population. The model parameters were estimated using least-squares methods based on dengue data in East Java, Indonesia. The model analysis results obtained two equilibria, namely the diseases-free and the endemic equilibria. The disease-free equilibrium is asymptotically stable if the basic reproduction number $ R_0 $ $ { < } $ 1, while the endemic equilibrium is asymptotically stable if $ R_0 $ $ { > } $ 1. The control variables were incorporated, and an optimal control problem was analyzed using the Pontryagin maximum principle. Finally, the cost-effectiveness analysis suggests that prevention only is the the most cost-effective strategy required to control dengue disease.
Citation: Anita T. Kurniawati, Fatmawati, Chidozie W. Chukwu, Windarto, Faishal F. Herdicho. Optimal control of dengue fever model with a logistically growing human population[J]. Mathematical Modelling and Control, 2025, 5(1): 48-60. doi: 10.3934/mmc.2025004
Dengue is a deadly illness in almost all parts of Indonesia, including East Java. This paper analyzes the dengue transmission model by considering the recruitment rate in the form of the logistic growth of the human population. The model parameters were estimated using least-squares methods based on dengue data in East Java, Indonesia. The model analysis results obtained two equilibria, namely the diseases-free and the endemic equilibria. The disease-free equilibrium is asymptotically stable if the basic reproduction number $ R_0 $ $ { < } $ 1, while the endemic equilibrium is asymptotically stable if $ R_0 $ $ { > } $ 1. The control variables were incorporated, and an optimal control problem was analyzed using the Pontryagin maximum principle. Finally, the cost-effectiveness analysis suggests that prevention only is the the most cost-effective strategy required to control dengue disease.

| [1] | World Health Organization, Dengue and severe dengue, 2023. |
| [2] | Ministry of Health of the Republic Indonesia, Hingga Juli Kasus DBD di Indonesia capai 71 ribu, 2020. |
| [3] |
M. Z. Ndii, Y. A. Adi, Understanding the effects of individual awareness and vector controls on malaria transmission dynamics using multiple optimal control, Chaos Solitions Fract., 153 (2021), 111476. https://doi.org/10.1016/j.chaos.2021.111476 doi: 10.1016/j.chaos.2021.111476

|
| [4] |
O. C. Collins, K. J. Duffy, A mathematical model for the dynamics and control of malaria in Nigeria, Infect. Dis. Model., 7 (2022), 728–741. https://doi.org/10.1016/j.idm.2022.10.005 doi: 10.1016/j.idm.2022.10.005
|
| [5] |
T. T. Yusuf, A. Abidemi, Effective strategies towards eradicating the tuberculosis epidemic: an optimal control theory alternative, Healthcare Anal., 3 (2023), 100131. https://doi.org/10.1016/j.health.2023.100193 doi: 10.1016/j.health.2023.100193
|
| [6] |
D. Aldila, B. L. Fardian, C. W. Chukwu, M. H. N. Aziz, P. Z. Kamalia, Improving tuberculosis control: assessing the value of medical masks and case detection–a multi-country study with cost-effectiveness analysis, R. Soc. Open Sci., 11 (2024), 231715. https://doi.org/10.1098/rsos.231715 doi: 10.1098/rsos.231715
|
| [7] |
O. J. Peter, N. D. Fahrani, Fatmawati, Windarto, C. W. Chukwu, A fractional derivative modeling study for measles infection with double dose vaccination, Healthcare Anal., 4 (2023), 100231. https://doi.org/10.1016/j.health.2023.100231 doi: 10.1016/j.health.2023.100231
|
| [8] |
M. V. Crankson, O. Olotu, A. S. Afolabi, A. Abidemi, Modeling the vaccination control of bacterial meningitis transmission dynamics: a case study, Math. Model. Control, 3 (2023), 416–434. https://doi.org/10.3934/mmc.2023033 doi: 10.3934/mmc.2023033
|
| [9] |
M. A. Khan, M. Z. Meetei, K. Shah, T. Abdeljawad, M. Y. Alshahrani, Modeling the monkeypox infection using the Mittag-Leffler kernel, Open Phys., 21 (2023), 20230111. https://doi.org/10.1515/phys-2023-0111 doi: 10.1515/phys-2023-0111
|
| [10] |
Z. U. A. Zafar, M. A. Khan, M. Inc, A. Akgül, M. Asiri, M. B. Riaz, The analysis of a new fractional model to the Zika virus infection with mutant, Heliyon, 10 (2024), e23390. https://doi.org/10.1016/j.heliyon.2023.e23390 doi: 10.1016/j.heliyon.2023.e23390
|
| [11] |
A. Abidemi, J. O. Akanni, O. D. Makinde, O. D. Makinde, A non-linear mathematical model for analysing the impact of COVID-19 disease on higher education in developing countries, Healthcare Anal., 3 (2023), 100193. https://doi.org/10.1016/j.health.2023.100193 doi: 10.1016/j.health.2023.100193
|
| [12] |
E. Alzahrani, M. A. Khan, Mathematical modeling and analysis of COVID‐19 infection: application to the Kingdom of Saudi Arabia data, J. Math., 2023 (2023), 6623005. https://doi.org/10.1155/2023/6623005 doi: 10.1155/2023/6623005
|
| [13] | E. Soewono, A. K. Supriatna, A two-dimentional model for the transmission of dengue fever disease, Bull. Malays. Math. Sci. Soc., 24 (2001), 49–57. |
| [14] |
A. Abdelrazec, J. Belair, C. Shan, H. Zhu, Modelling the spread and control of dengue with limited public health resources, Math. Biosci., 271 (2016), 136–145. https://doi.org/10.1016/j.mbs.2015.11.004 doi: 10.1016/j.mbs.2015.11.004
|
| [15] |
L. Esteva, H. M. Yang, Assessing the effects of temperature and dengue virus load on dengue transmission, J. Biol. Syst., 23 (2015), 573–554. https://doi.org/10.1142/S0218339015500278 doi: 10.1142/S0218339015500278
|
| [16] |
R. Taghikhani, A. B. Gumel, Mathematics of dengue transmission dynamics: roles of vector vertical transmission and temperature fluctuations, Infect. Dis. Model., 3 (2018), 266–292. https://doi.org/ 10.1016/j.idm.2018.09.003 doi: 10.1016/j.idm.2018.09.003
|
| [17] |
N. Anggriani, H. Tasman, M. Z. Ndii, A. K. Supriatna, E. Soewono, E. Siregar, The effect of reinfection with the same serotype on dengue transmission dynamics, Appl. Math. Comput., 349 (2019), 62–80. https://doi.org/10.1016/j.amc.2018.12.022 doi: 10.1016/j.amc.2018.12.022
|
| [18] |
E. Bonyah, M. L. Juga, C. W. Chukwu, Fatmawati, A fractional order dengue fever model in the context of protected travellers, Alex. Eng. J., 61 (2022), 927–936. https://doi.org/10.1016/j.aej.2021.04.070 doi: 10.1016/j.aej.2021.04.070
|
| [19] |
Windarto, M. A. Khan, Fatmawati, Parameter estimation and fractional derivatives of dengue transmission model, AIMS Math., 5 (2020), 2758–2779. https://doi.org/10.3934/math.2020178 doi: 10.3934/math.2020178
|
| [20] |
M. A. Khan, Fatmawati, Dengue infection modeling and its optimal control analysis in East Java, Indonesia, Heliyon, 7 (2021), e06023. https://doi.org/10.1016/j.heliyon.2021.e06023 doi: 10.1016/j.heliyon.2021.e06023
|
| [21] |
M. Z. Ndii, The effects of vaccination, vector controls and media on dengue transmission dynamics with a seasonally varying mosquito population, Results Phys., 34 (2022), 105298. https://doi.org/10.1016/j.rinp.2022.105298 doi: 10.1016/j.rinp.2022.105298
|
| [22] |
J. Zhang, L. Liu, Y. Li, Y. Wang, An optimal control problem for dengue transmission model with Wolbachia and vaccination, Commun. Nonlinear Sci. Numer. Simul., 116 (2023), 106856. https://doi.org/10.1016/j.cnsns.2022.106856 doi: 10.1016/j.cnsns.2022.106856
|
| [23] |
A. T. Kurniawati, Fatmawati, Windarto, Global analysis of a dengue hemorrhagic fever transmission model with logistics growth in human population, AIP Conf. Proc., 2329 (2021), 040007. https://doi.org/10.1063/5.0042364 doi: 10.1063/5.0042364
|
| [24] |
M. Samsuzzoha, M. Singh, D. Lucy, Parameter estimation of influenza epidemic model, Appl. Math. Comput., 220 (2013), 616–629. https://doi.org/10.1016/j.amc.2013.07.040 doi: 10.1016/j.amc.2013.07.040
|
| [25] |
Fatmawati, C. W. Chukwu, R. T. Alqahtani, C. Alfiniyah, F. F. Herdicho, Tasmi, A Pontryagin's maximum principle and optimal control model with cost-effectiveness analysis of the COVID-19 epidemic, Decis. Anal. J., 8 (2023), 100273. https://doi.org/10.1016/j.dajour.2023.100273 doi: 10.1016/j.dajour.2023.100273
|
| [26] | Health office (Dinas Kesehatan) of East Java province, Data of dengue, 2021. |
| [27] | Central Bureau of Statistics East Java Province, Angka harapan hidup (Tahun), 2017–2019, 2023. |
| [28] | Central Bureau of Statistics East Java Province, Angka harapan hidup (Tahun), 2020–2022, 2023. |
| [29] | I. Mahmood, M. Jahan, D. Groen, A. Javed, F. Shafait, An agent-based simulation of the spread of dengue fever, In: V. V. Krzhizhanovskaya, G. Závodszky, M. H. Lees, J. J. Dongarra, P. M. A. Sloot, S. Brissos, et al., Computational Science-ICCS 2020, 2020,103–117. https://doi.org/10.1007/978-3-030-50420-5_8 |
| [30] | Central Bureau of Statistics East Java Province, Jumlah penduduk menurut jenis kelamin dan Kabupaten/Kota Provinsi Jawatimur (Jiwa), 2018–2020, 2023. |
| [31] |
P. van den Driessche, J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci. 180 (2002), 29–48. https://doi.org/10.1016/S0025-5564(02)00108-6 doi: 10.1016/S0025-5564(02)00108-6
|
| [32] |
N. Chitnis, J. M. Hyman, J. M. Cushing, Determining important parameters in the spread of malaria through the sensitivity analysis of a mathematical model, Bull. Math. Biol., 70 (2008), 1272–1296. https://doi.org/10.1007/s11538-008-9299-0 doi: 10.1007/s11538-008-9299-0
|
| [33] |
F. Agusto, M. A. Khan, Optimal control strategies for dengue transmission in Pakistan, Math. Biosci., 305 (2018), 102–121. https://doi.org/10.1016/j.mbs.2018.09.007 doi: 10.1016/j.mbs.2018.09.007
|
| [34] |
S. Ullah, M. F. Khan, S. A. A. Shah, M. Farooq, M. A. Khan, M. bin Mamat, Optimal control analysis of vector-host model with saturated treatment, Eur. Phys. J. Plus, 135 (2020), 839. https://doi.org/10.1140/epjp/s13360-020-00855-1 doi: 10.1140/epjp/s13360-020-00855-1
|
| [35] | L. S. Pontryagin, The mathematical theory of optimal processes, CRC Press, 1987. https://doi.org/10.1201/9780203749319 |
| [36] | S. Lenhart, J. T. Workman, Optimal control applied to biological models, Chapman and Hall/CRC, 2007. https://doi.org/10.1201/9781420011418 |
| [37] |
B. Buonomo, R. D. Marca, Optimal bed net use for a dengue disease model with mosquito seasonal pattern, Math. Methods Appl. Sci., 41 (2017), 573–592. https://doi.org/10.1002/mma.4629 doi: 10.1002/mma.4629
|
| [38] |
J. K. K. Asamoah, E. Yankson, E. Okyere, G. G. Sun, Z. Jin, R. Jan, et al., Optimal control and cost-effectiveness analysis for dengue fever model with asymptomatic and partial immune individuals, Results Phys., 31 (2021), 104919. https://doi.org/10.1016/j.rinp.2021.104919 doi: 10.1016/j.rinp.2021.104919
|










Figures(11) / Tables(7)
Anita T. Kurniawati, Fatmawati, Chidozie W. Chukwu, Windarto, Faishal F. Herdicho. Optimal control of dengue fever model with a logistically growing human population[J]. Mathematical Modelling and Control, 2025, 5(1): 48-60. doi: 10.3934/mmc.2025004







 DownLoad:
DownLoad: