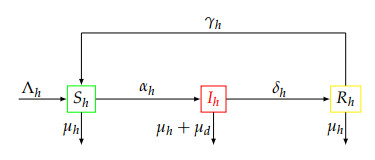
A deterministic multi-stage malaria model with a non-therapeutic control measure and the effect of loss of immunity due to the use of the Long-Lasting bednets with a control perspective is formulated and analyzed both theoretically and numerically. The model basic reproduction number is derived, and analytical results show that the model's equilibria are locally and globally asymptotically stable when certain threshold conditions are satisfied. Pontryagin's Maximum Principle with respect to a time dependent constant is used to derive the necessary conditions for the optimal usage of the Long-Lasting Insecticide-treated bednets (LLINs) to mitigate the malaria transmission dynamics. This is accomplished by introducing biologically admissible controls and $ \epsilon\% $-approximate sub-optimal controls. Forward-backward fourth-order Runge-Kutta method is used to numerically solve the optimal control problem. We observe that the disadvantage (loss of immunity, even at its maximum) in the use of bednets is compensated by the benefit of the number of susceptible/infected individuals excluded from the malaria disease dynamics, the only danger being the poor use of the long-lasting bednets. Moreover, it is possible to get closer to the optimal results with a realistic strategy. The results from this study could help public health planners and policy decision-makers to design reachable and more practical malaria prevention programs "close" to the optimal strategy.
Citation: S. Y. Tchoumi, Y. Kouakep-Tchaptchie, D. J. Fotsa-Mbogne, J. C. Kamgang, J. M. Tchuenche. Optimal control of a malaria model with long-lasting insecticide-treated nets[J]. Mathematical Modelling and Control, 2021, 1(4): 188-207. doi: 10.3934/mmc.2021018
A deterministic multi-stage malaria model with a non-therapeutic control measure and the effect of loss of immunity due to the use of the Long-Lasting bednets with a control perspective is formulated and analyzed both theoretically and numerically. The model basic reproduction number is derived, and analytical results show that the model's equilibria are locally and globally asymptotically stable when certain threshold conditions are satisfied. Pontryagin's Maximum Principle with respect to a time dependent constant is used to derive the necessary conditions for the optimal usage of the Long-Lasting Insecticide-treated bednets (LLINs) to mitigate the malaria transmission dynamics. This is accomplished by introducing biologically admissible controls and $ \epsilon\% $-approximate sub-optimal controls. Forward-backward fourth-order Runge-Kutta method is used to numerically solve the optimal control problem. We observe that the disadvantage (loss of immunity, even at its maximum) in the use of bednets is compensated by the benefit of the number of susceptible/infected individuals excluded from the malaria disease dynamics, the only danger being the poor use of the long-lasting bednets. Moreover, it is possible to get closer to the optimal results with a realistic strategy. The results from this study could help public health planners and policy decision-makers to design reachable and more practical malaria prevention programs "close" to the optimal strategy.

| [1] | H. Abboubakar, R. Racke, Mathematical modelling and optimal control of typhoid fever, Konstanzer Schriften in Mathematik, 386 (2019), 1–32. |
| [2] | A. I. Abioye, O. J. Peter, A. A. Ayoade, O. A. Uwaheren, M. O. Ibrahim, Application of adomian decomposition method on a mathematical model of malaria, Advances in Mathematics: Scientific Journal, 9 (2020). |
| [3] | A. I. Abioye, O. J. Peter, F. A. Oguntolu, A. F. Adebisi, T. F. Aminu, Global stability of seir-sei model of malaria transmission, Adv. Math., Sci. J, 9 (2020). 5305–5317. |
| [4] | A. I. Abioye, M. O. Ibrahim, O. J. Peter, H. A. Ogunseye, Optimal control on a mathematical model of malaria, Sci. Bull., Series A: Appl Math Phy, (2020), 178–190. |
| [5] | AI Abioye, MO Ibrahim, OJ Peter, S Amadiegwu, FA Oguntolu, Differential transform method for solving mathematical model of seir and sei spread of malaria, 2018. |
| [6] |
F. B. Agusto, S. Y. Del Valle, K. W. Blayneh, C. N. Ngonghala, M. J. Goncalves, N. Li, et al., The impact of bed-net use on malaria prevalence, J. Theor.Biol., 320 (2013), 58–65. doi: 10.1016/j.jtbi.2012.12.007

|
| [7] | M. E. Alexander, S. M. Moghadas, P. Rohani, A. R. Summers. Modelling the effect of a booster vaccination on disease epidemiology, J. Math. Biol., (2006), 290–306. |
| [8] |
R. Anguelov, Y. Dumont, J. Lubuma, E. Mureithi, Stability analysis and dynamics preserving nonstandard finite difference schemes for a malaria model, Mathematical Population Studies: An International Journal of Mathematical Demography, 20 (2013), 101–122. doi: 10.1080/08898480.2013.777240
|
| [9] | J. Arino, K. L. Cooke, P. van den Driessche, J. Velasco-Hernández, An epidemiology model that includes a leaky vaccine with a general waning function, Discrete Contin. Dyn. Syst. Ser. B, 4 (2004), 479–495. |
| [10] | O. J. Briet, N. Chitnis, Effects of changing mosquito host searching behaviour on the cost effectiveness of a mass distribution of long-lasting, insecticidal nets: A modelling study, Malaria J., 12 (2013). |
| [11] | B. Buonomo, Analysis of a malaria model with mosquito host choice and bed-net control, Int. J. Biomath., 8 (2015). |
| [12] | N. Chitnis, Using Mathematical models in controlling the spread of malaria, PhD thesis, University of Arizona, 2005. |
| [13] | N. Chitnis, J. M. Hyman, J. Cushing, Determining Important Parameters in the Spread of Malaria Through the Sensitivity Analysis of a Mathematical Model, Bull. Math. Biol., 2008. |
| [14] |
J. Dushoff, W. Huang, C. Castillo-Chavez, Backwards bifurcations and catastrophe in simple models of fatal diseases, J. Math. Biol., 36 (1998), 227–248. doi: 10.1007/s002850050099
|
| [15] | D. Gollin, C. Zimmermann, Malaria: Disease impacts and long-run income differences, IZA Discussion Papers 2997, Institution for the Study of Labor (IZA), 2007. |
| [16] |
C. A. Guerra, A. M. Noor, H. Y. Myint, S. I. Hay, R. W. Snow, The global distribution of clinical episodes of plasmodium falciparum malaria, Nature, 434 (2005), 214–217. doi: 10.1038/nature03342
|
| [17] | J. C. Kamgang, V. C. Kamla, S. Y. Tchoumi. Modeling the dynamic of transmission of the malaria with bed net protection perspective, SIAM J. Appl. Math., 2014. |
| [18] |
J. C. Kamgang, G. Sallet, Computation of threshold conditions for epidemiological models and global stability of the disease free equilibrium, Math. Biosci., 213 (2008), 1–12. doi: 10.1016/j.mbs.2008.02.005
|
| [19] | J. C. Kamgang, C. P. Thron, Analysis of malaria control measures' effectiveness using multistage vector model, Bulletin of Mathematical Biology, 2019. |
| [20] | J. C. Kamgang, S. Y. Tchoumi, A model of the dynamic of transmission of malaria, integrating seirs, seis, sirs and sis organization in the host–population, J. Appl. Anal. Comput., 5 (2015). |
| [21] | C. M. Kribs-Zaleta, J. X. Velasco-Hernández, A simple vaccination model with multiple endemic states, Math. Biosci., 8 (2000), 183–201. |
| [22] | A. Ali Lashari, K. Hattaf, G. Zaman, X.-Z. Li, Backward bifurcation and optimal control of a vector borne disease, Applied Mathematics & Information Sciences, 7 (2013), 301–309. |
| [23] |
A. Ali Lashari, G. Zaman, Optimal control of a vector borne disease with horizontal transmission, Nonlinear Analysis: Real World Applications, 13 (2012), 203–212. doi: 10.1016/j.nonrwa.2011.07.026
|
| [24] | S. Lenhart, Optimal control theory in application to biology. Short course lectures and lab notes from 2003, University of Tennessee, Knoxville, 2003. |
| [25] | S. Lenhart, J. T. Workman, Optimal control applied to biological models, Chapman and Hall/CRC, 2007. |
| [26] | G. Macdonald, The epidemiology and control of malaria, London, Oxford University Press, 1957. |
| [27] | Moiroux, Modélisation du risque d'exposition aux moustiques vecteurs du plas- modium spp. dans un contexte de lutte anti-vectorielle. Ecologie, environnement. Université de Montpellier II, 2012. |
| [28] | C. Ndo, B. Menze-Djantio, C. Antonio-Nkondjio, Awareness, attitudes and prevention of malaria in the cities of douala and yaoundé (cameroon), Parasites and Vectors, 181 (2011). |
| [29] | S. Olaniyi, OS Obabiyi, Mathematical model for malaria transmission dynamics in human and mosquito populations with nonlinear forces of infection, Int. J. Pure Appl. Math., 88 (2013), 125–156. |
| [30] | SA Pedro, H Rwezaura, JM Tchuenche, Time-varying sensitivity analysis of an influenza model with interventions, Int. J. Biomath., (2021), 2150098. |
| [31] | L. Pontryagin, V. Boltyanskii, R. Gamkrelidze, E. Mishchenko, The mathematical theory of optimal control process, 1986. |
| [32] |
A.-M. Pulkki-Brannstrom, C. Wolff, N. Brannstrom, J. Skordis-Worrall, Cost and cost effectiveness of long-lasting insecticide-treated bed nets - a model-based analysis, Cost Effectiveness and Resource Allocation, 10 (2012), 5. doi: 10.1186/1478-7547-10-5
|
| [33] | P. Resseguier, Contribution à l'étude du repas sanguin de culex pipiens pipiens, PhD thesis, Université de Toulouse, 2011. |
| [34] |
V. Robert, K. Macintyre, J. Keating, J. F. Trape, J. B. Duchemin, M. Warren, et al., Malaria transmission in urban sub- saharan africa, Am. J. Trop. Med. Hyg., 68 (2003), 169 –176. doi: 10.4269/ajtmh.2003.68.169
|
| [35] | R. Ross, The prevention of malaria, John Murray, 1911. |
| [36] |
R. Short, R. Gurung, M. Rowcliffe, N. Hill, E. J. Milner-Gulland, The use of mosquito nets in fisheries: A global perspective, PLoS ONE, 13 (2018), e0191519. doi: 10.1371/journal.pone.0191519
|
| [37] | S. Y. Tchoumi, J. C. Kamgang, D. Tieudjo, G. Sallet, A basic general model of vector-borne diseases, Commun. Math. Biol. Neurosci, 3 (2018). |
| [38] | J. M. Tchuenche, C. Chiyaka, D. Chan, A. Matthews, G. Mayer, A mathematical model for antimalarial drug resistance, Mathematical Medicine and Biology, (2010), 1–21. |
| [39] | WHO, www.who.int. Technical report, WHO, 2018. |














Figures(15) / Tables(4)
S. Y. Tchoumi, Y. Kouakep-Tchaptchie, D. J. Fotsa-Mbogne, J. C. Kamgang, J. M. Tchuenche. Optimal control of a malaria model with long-lasting insecticide-treated nets[J]. Mathematical Modelling and Control, 2021, 1(4): 188-207. doi: 10.3934/mmc.2021018







 DownLoad:
DownLoad: