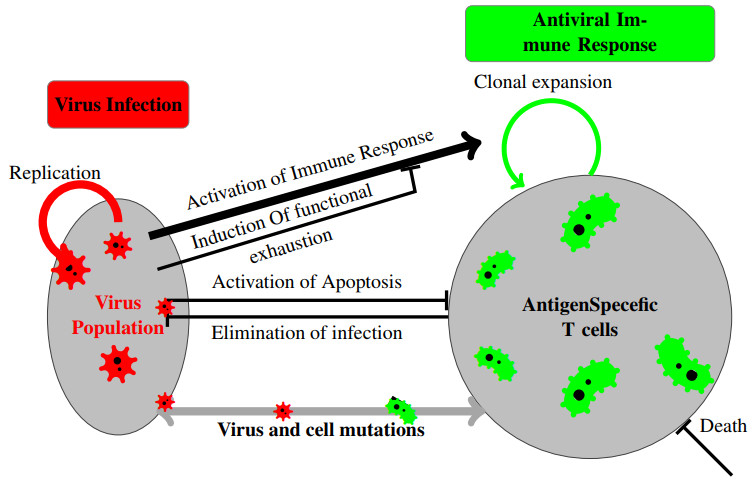
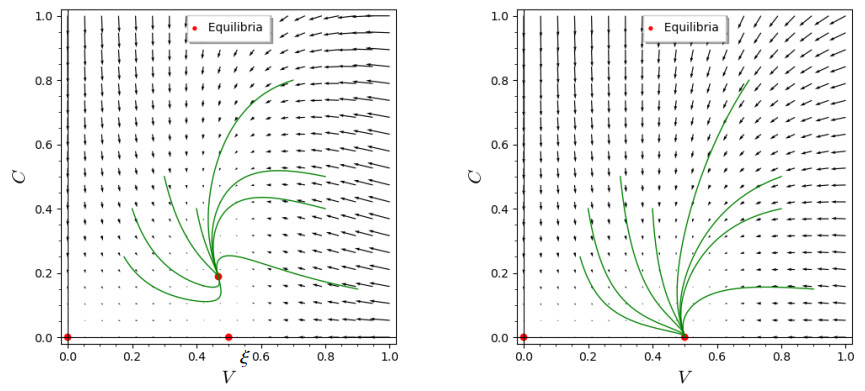
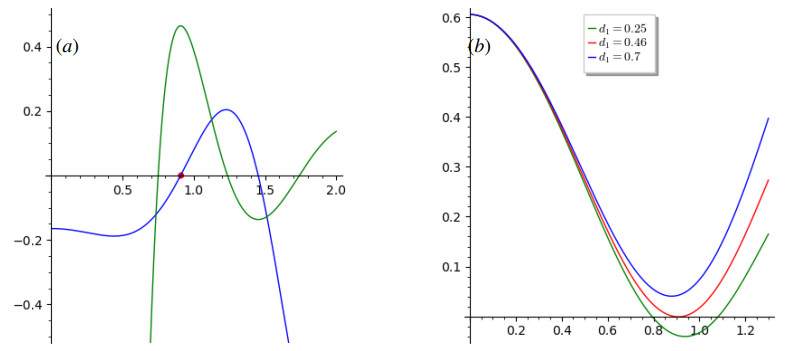
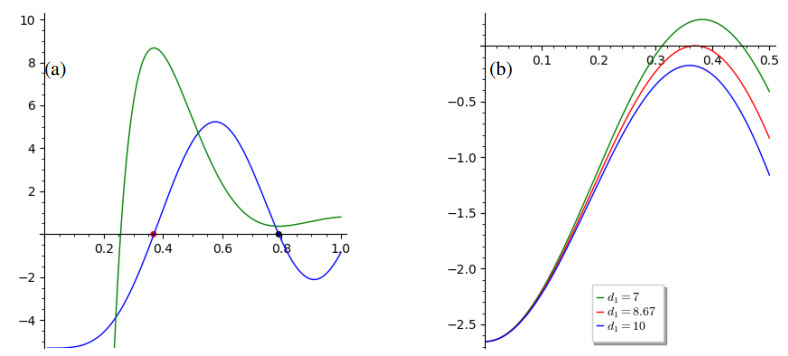
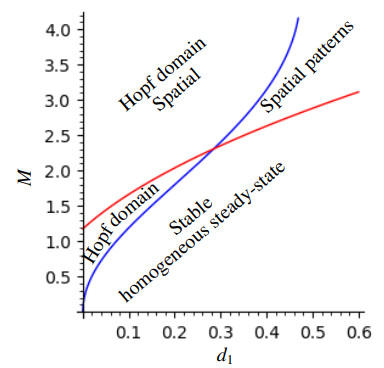
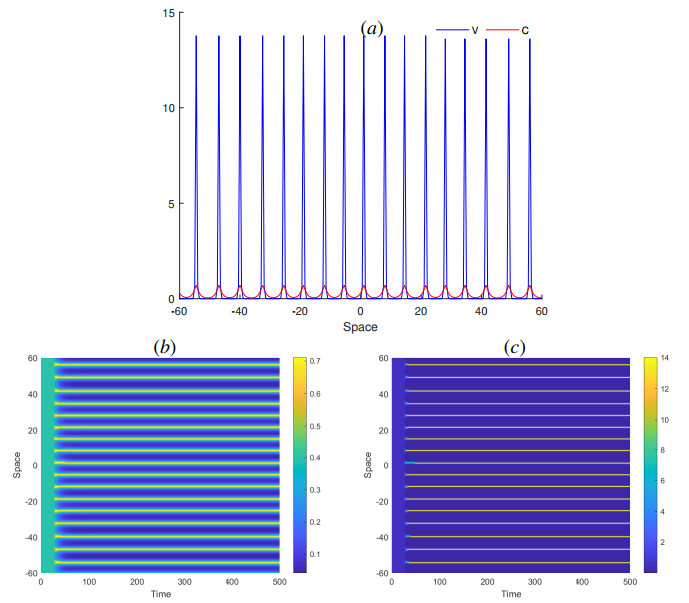
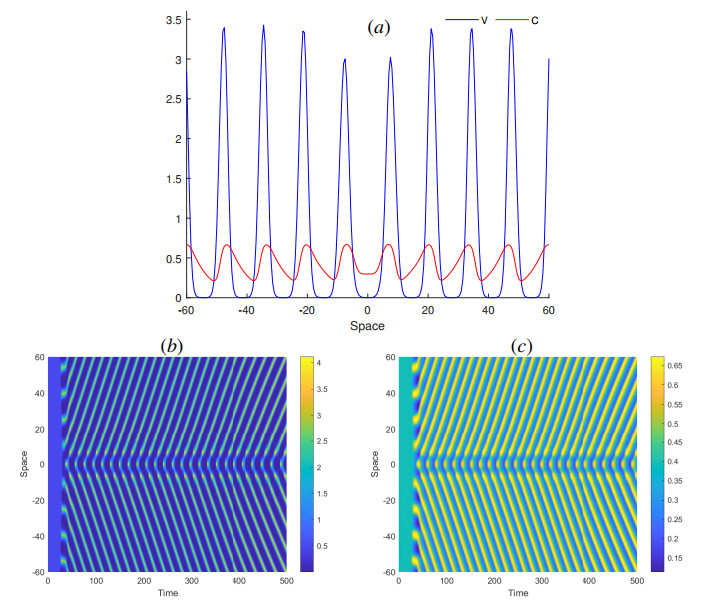
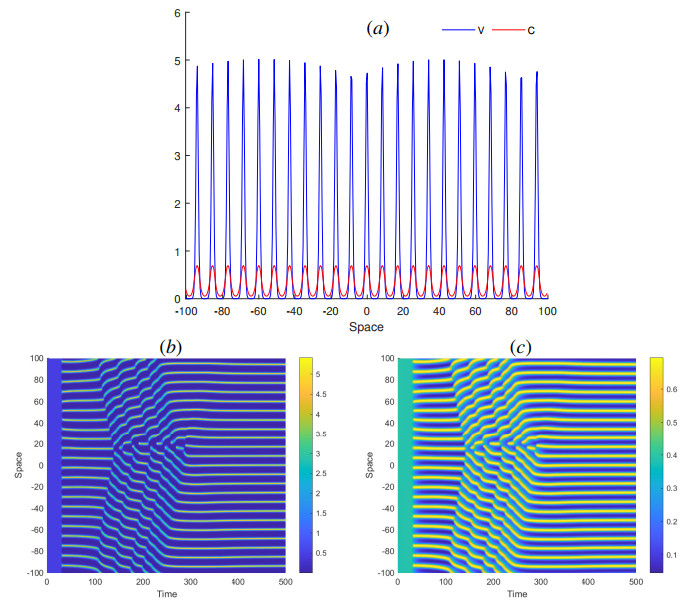
The process of viral infection spreading in tissues was influenced by various factors, including virus replication within host cells, transportation, and the immune response. Reaction-diffusion systems provided a suitable framework for examining this process. In this work, we studied a nonlocal reaction-diffusion system of equations that modeled the distribution of viruses based on their genotypes and their interaction with the immune response. It was shown that the infection may persist at a certain level alongside a chronic immune response, exhibiting spatially uniform or oscillatory behavior. Finally, the immune cells may become entirely depleted, leading to a high viral load persisting in the tissue. Numerical simulations were employed to elucidate the nonlinear dynamics and pattern formation inherent in the nonlocal model.
Citation: Ali Moussaoui, Vitaly Volpert. The impact of immune cell interactions on virus quasi-species formation[J]. Mathematical Biosciences and Engineering, 2024, 21(11): 7530-7553. doi: 10.3934/mbe.2024331
The process of viral infection spreading in tissues was influenced by various factors, including virus replication within host cells, transportation, and the immune response. Reaction-diffusion systems provided a suitable framework for examining this process. In this work, we studied a nonlocal reaction-diffusion system of equations that modeled the distribution of viruses based on their genotypes and their interaction with the immune response. It was shown that the infection may persist at a certain level alongside a chronic immune response, exhibiting spatially uniform or oscillatory behavior. Finally, the immune cells may become entirely depleted, leading to a high viral load persisting in the tissue. Numerical simulations were employed to elucidate the nonlinear dynamics and pattern formation inherent in the nonlocal model.

| [1] |
P. Auger, A. Moussaoui, On the threshold of release of confinement in an epidemic SEIR model taking into account the protective effect of mask, Bull. Math. Biol., 83 (2021), 25. https://doi.org/10.1007/s11538-021-00858-8 doi: 10.1007/s11538-021-00858-8

|
| [2] |
A. Moussaoui, M. Meziane, On the date of the epidemic peak, Math. Biosci. Eng., 21 (2024), 2835–2855. https://doi.org/10.3934/mbe.2024126 doi: 10.3934/mbe.2024126
|
| [3] |
A. Moussaoui, E. H. Zerga, Transmission dynamics of COVID-19 in Algeria: The impact of physical distancing and face masks, AIMS Public Health, 7 (2020), 816–827. https://doi.org/10.3934/publichealth.2020063 doi: 10.3934/publichealth.2020063
|
| [4] |
A. Moussaoui, P. Auger, Prediction of confinement effects on the number of COVID-19 outbreaks in Algeria, Math. Model. Nat. Phenom., 15 (2020), 37. https://doi.org/10.1051/mmnp/2020028 doi: 10.1051/mmnp/2020028
|
| [5] | A. Moussaoui, Estimating the final size relation for the SIR discrete epidemic model under intervention, Nonlinear Stud., 31 (2024), 1. https://nonlinearstudies.com/index.php/nonlinear/article/view/3221 |
| [6] |
T. Nguyen-Huu, P. Auger, A. Moussaoui, On incidence-dependent management strategies against an SEIRS epidemic: Extinction of the epidemic using Allee effect, Mathematics, 11 (2023), 2822. https://doi.org/10.3390/math11132822 doi: 10.3390/math11132822
|
| [7] |
G. Bocharov, A. Meyerhans, N. Bessonov, S. Trofimchuk, V. Volpert, Interplay between reaction and diffusion processes in governing the dynamics of virus infections, J. Theor. Biol., 457 (2018), 221–236. https://doi.org/10.1016/j.jtbi.2018.08.036. doi: 10.1016/j.jtbi.2018.08.036
|
| [8] |
J. N. Mandl, P. Torabi-Parizi, R. N. Germain, Visualization and dynamic analysis of host-pathogen interactions, Curr. Opin. Immunol., 29 (2014), 8–15. https://doi.org/10.1016/j.coi.2014.03.002 doi: 10.1016/j.coi.2014.03.002
|
| [9] |
X. Sewald, N. Motamedi, W. Mothes, Viruses exploit the tissue physiology of the host to spread in vivo, Curr. Opin. Cell Biol., 41 (2016), 81–90. https://doi.org/10.1016/j.ceb.2016.04.008 doi: 10.1016/j.ceb.2016.04.008
|
| [10] | C. K. Biebricher, M. Eigen, What is a quasispecies?, Curr. Top Microbiol. Immunol., 299 (2006), 1–31. |
| [11] |
E. Domingo, J. Sheldon, C. Perales, Viral quasispecies evolution, Microbiol. Mol. Biol. Rev., 76 (2012), 159–216. https://doi.org/10.1128/mmbr.05023-11 doi: 10.1128/mmbr.05023-11
|
| [12] | M. Nowak, R. M. May, Virus Dynamics, in Mathematical Principles of Immunology and Virology, Oxford University Press, 2000. |
| [13] |
M. Meziane, A. Moussaoui, V. Volpert, On a two-strain epidemic model involving delay equations, Math. Biosci. Eng., 20 (2023), 20683–20711. https://doi.org/10.3934/mbe.2023915 doi: 10.3934/mbe.2023915
|
| [14] |
N. Bessonov, G. Bocharov, A. Meyerhans, V. Popov, V. Volpert, Existence and dynamics of strains in a nonlocal reaction-diffusion model of viral evolution, SIAM J. Appl. Math., 81 (2021), 107–128, https://doi.org/10.1137/19M1282234 doi: 10.1137/19M1282234
|
| [15] |
M. Kimura, Diffusion models in population genetics, J. Appl. Probab., 1 (1964), 177–232. https://doi.org/10.2307/3211856 doi: 10.2307/3211856
|
| [16] |
A. Sasaki, Evolution of antigen drift/switching: continuously evading pathogens, J. Theor. Biol., 163 (1994), 291–308. https://doi.org/10.1006/jtbi.1994.1110 doi: 10.1006/jtbi.1994.1110
|
| [17] |
N. Bessonov, G. A. Bocharov, C. Leon, V. Popov, V. Volpert, Genotype-dependent virus distribution and competition of virus strains, Math. Mech. Complex Syst., 8 (2020), 101–126. https://doi.org/10.2140/memocs.2020.8.101 doi: 10.2140/memocs.2020.8.101
|
| [18] |
A. Moussaoui, V. Volpert, The influence of immune cells on the existence of virus quasi-species, Math. Biosci. Eng., 20 (2023), 15942–15961. https://doi.org/10.3934/mbe.2023710 doi: 10.3934/mbe.2023710
|
| [19] |
G. Bocharov, A. Meyerhans, N. Bessonov, S. Trofimchuk, V. Volpert, Modelling the dynamics of virus infection and immune response in space and time, Int. J. Parallel, Emergent Distrib. Syst., 34 (2019), 341–355. https://doi.org/10.1080/17445760.2017.1363203 doi: 10.1080/17445760.2017.1363203
|
| [20] |
A. Mozokhina, L. A. Mahiout, V. Volpert, Modeling of viral infection with inflammation, Mathematics, 11 (2023), 4095. https://doi.org/10.3390/math11194095 doi: 10.3390/math11194095
|
| [21] | V. Volpert, Elliptic Partial Differential Equations, Volume 2: Reaction-Diffusion Equations, Villeurbanne: Birkhäuser, 2014. |
| [22] | A. Volpert, V. Volpert, Spectrum of elliptic operators and stability of travelling waves, Asymptotic Anal., 23 (2000), 111–134. |
| [23] |
B. Anderson, J. Jackson, M. Sitharam, Descartes rule of signs revisited, Am. Math. Mon., 105 (1998), 447–151. https://doi.org/10.1080/00029890.1998.12004907 doi: 10.1080/00029890.1998.12004907
|
| [24] | J. D. Murray, Mathematical Biology II, Heidelberg, Springer-Verlag, 2002. |
| [25] | R. S. Strichartz, A Guide to Distribution Theory and Fourier Transforms, World Scientific, Singapour, 2003. |
| [26] |
S. Genieys, V. Volpert, P. Auger, Pattern and waves for a model in population dynamics with nonlocal consumption of resources, Math. Model. Nat. Phenom., 1 (2006), 63–80. https://doi.org/10.1051/mmnp:2006004 doi: 10.1051/mmnp:2006004
|
| [27] |
B. L Segal, V. Volpert, A. Bayliss, Pattern formation in a model of competing populations with nonlocal interactions, Phys. D: Nonlinear Phenom., 253 (2013), 12–23. https://doi.org/10.1016/j.physd.2013.02.006 doi: 10.1016/j.physd.2013.02.006
|
| [28] |
N. Bessonov, N. Reinberg, M. Banerjee, V. Volpert, The origin of species by means of mathematical modelling, Acta Biotheor., 66 (2018), 333–344. https://doi.org/10.1007/s10441-018-9328-9 doi: 10.1007/s10441-018-9328-9
|
| [29] |
N. Bessonov, D. Neverova, V. Popov, V. Volpert, Emergence and competition of virus variants in respiratory viral infections, Front. Immunol., 13 (2023), 945228. https://doi.org/10.3389/fimmu.2022.945228. doi: 10.3389/fimmu.2022.945228
|
Figures(8)

Ali Moussaoui, Vitaly Volpert. The impact of immune cell interactions on virus quasi-species formation[J]. Mathematical Biosciences and Engineering, 2024, 21(11): 7530-7553. doi: 10.3934/mbe.2024331







 DownLoad:
DownLoad: