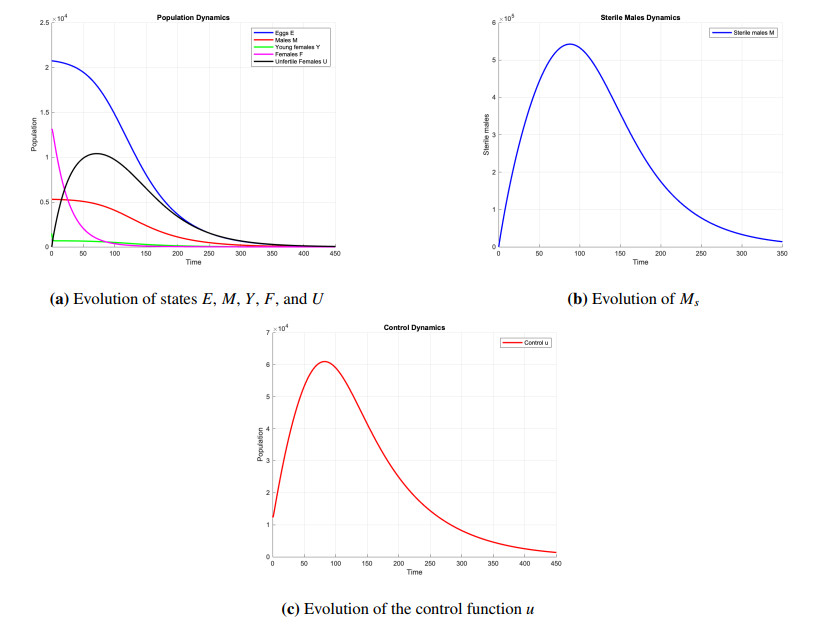
This paper focuses on the feedback global stabilization and observer construction for a sterile insect technique model. The sterile insect technique (SIT) is one of the most ecological methods for controlling insect pests responsible for worldwide crop destruction and disease transmission. In this work, we construct a feedback law that globally asymptotically stabilizes an SIT model at extinction equilibrium. Since the application of this type of control requires the measurement of different states of the target insect population, and, in practice, some states are more difficult or more expensive to measure than others, it is important to know how to construct a state estimator, which from a few well-chosen measured states, estimates the other ones, as the one we build in the second part of our work. In the last part of our work, we show that we can apply the feedback control with estimated states to stabilize the full system.
Citation: Kala Agbo Bidi. Feedback stabilization and observer design for sterile insect technique models[J]. Mathematical Biosciences and Engineering, 2024, 21(6): 6263-6288. doi: 10.3934/mbe.2024274
This paper focuses on the feedback global stabilization and observer construction for a sterile insect technique model. The sterile insect technique (SIT) is one of the most ecological methods for controlling insect pests responsible for worldwide crop destruction and disease transmission. In this work, we construct a feedback law that globally asymptotically stabilizes an SIT model at extinction equilibrium. Since the application of this type of control requires the measurement of different states of the target insect population, and, in practice, some states are more difficult or more expensive to measure than others, it is important to know how to construct a state estimator, which from a few well-chosen measured states, estimates the other ones, as the one we build in the second part of our work. In the last part of our work, we show that we can apply the feedback control with estimated states to stabilize the full system.

| [1] |
H. Barclay, M. Mackauer, The sterile insect release method for pest control: A density-dependent model, Environ. Entomol., 9 (1980), 810–817. https://doi.org/10.1093/ee/9.6.810 doi: 10.1093/ee/9.6.810

|
| [2] |
R. Gato, Z. Menéndez, E. Prieto, R. Argilés, M. Rodríguez, W. Baldoquín, et al., Sterile insect technique: Successful suppression of an aedes aegypti field population in Cuba, Insects, 12 (2021), 469. https://doi.org/10.3390/insects12050469 doi: 10.3390/insects12050469
|
| [3] | M. J. Vreysen, J. Hendrichs, W. R. Enkerlin, The sterile insect technique as a component of sustainable area-wide integrated pest management of selected horticultural insect pests, J. Fruit Ornamental Plant Res., 14 (2006), 107. |
| [4] |
M. Strugarek, H. Bossin, Y. Dumont, On the use of the sterile insect release technique to reduce or eliminate mosquito populations, Appl. Math. Modell., 68 (2019), 443–470. https://doi.org/10.1016/j.apm.2018.11.026 doi: 10.1016/j.apm.2018.11.026
|
| [5] |
L. Almeida, M. Duprez, Y. Privat, N. Vauchelet, Optimal control strategies for the sterile mosquitoes technique, J. Differ. Equations, 311 (2022), 229–266. https://doi.org/10.1016/j.jde.2021.12.002 doi: 10.1016/j.jde.2021.12.002
|
| [6] |
L. Almeida, J. Estrada, N. Vauchelet, Wave blocking in a bistable system by local introduction of a population: Application to sterile insect techniques on mosquito populations, Math. Modell. Nat. Phenom., 17 (2022), 22, https://doi.org/10.1051/mmnp/2022026 doi: 10.1051/mmnp/2022026
|
| [7] |
L. Almeida, A. Léculier, N. Vauchelet, Analysis of the rolling carpet strategy to eradicate an invasive species, SIAM J. Math. Anal., 55 (2023), 275–309. https://doi.org/10.1137/21M1427243 doi: 10.1137/21M1427243
|
| [8] |
A. Leculier, N. Nguyen, A control strategy for the sterile insect technique using exponentially decreasing releases to avoid the hair-trigger effect, Math. Model. Nat. Phenom., 18 (2023), 25. https://doi.org/10.1051/mmnp/2023018 doi: 10.1051/mmnp/2023018
|
| [9] |
P. A. Bliman, Y. Dumont, Robust control strategy by the sterile insect technique for reducing epidemiological risk in presence of vector migration, Math. Biosci., 350 (2022), 108856. https://doi.org/10.1016/j.mbs.2022.108856 doi: 10.1016/j.mbs.2022.108856
|
| [10] |
P. A. Bliman, D. Cardona-Salgado, Y. Dumont, O. Vasilieva, Implementation of control strategies for sterile insect techniques, Math. Biosci., 314 (2019), 43–60. https://doi.org/10.1016/j.mbs.2019.06.002 doi: 10.1016/j.mbs.2019.06.002
|
| [11] | K. A. Bidi, L. Almeida, J. M. Coron, Global stabilization of sterile insect technique model by feedback laws, preprint, arXiv: 2307.00846. https://doi.org/10.48550/arXiv.2307.00846 |
| [12] |
R. E. Kalman, A new approach to linear filtering and prediction problems, J. Basic Eng., 82 (1960), 35–45. https://doi.org/10.1115/1.3662552 doi: 10.1115/1.3662552
|
| [13] |
D. G. Luenberger, Observing the state of a linear system, IEEE Trans. Mil. Electron., 8 (1964), 74–80. https://doi.org/10.1109/TME.1964.4323124 doi: 10.1109/TME.1964.4323124
|
| [14] |
R. Anguelov, Y. Dumont, J. Lubuma, Mathematical modeling of sterile insect technology for control of anopheles mosquito, Comput. Math. Appl., 64 (2012), 374–389. https://doi.org/10.1016/j.camwa.2012.02.068 doi: 10.1016/j.camwa.2012.02.068
|
| [15] |
L. Esteva, H. M. Yang, Mathematical model to assess the control of aedes aegypti mosquitoes by the sterile insect technique, Math. Biosci., 198 (2005), 132–147. https://doi.org/10.1016/j.mbs.2005.06.004 doi: 10.1016/j.mbs.2005.06.004
|
| [16] | P. A. Bliman, Feedback control principles for biological control of dengue vectors, in 2019 18th European Control Conference (ECC), (2019), 1659–1664. https://doi.org/10.23919/ECC.2019.8795984 |
| [17] | A. Cristofaro, L. Rossi, Backstepping control for the sterile mosquitoes technique: Stabilization of extinction equilibrium, unpublished work. |
| [18] | A. F. Filippov, Differential equations with discontinuous right-hand side, Mat. Sb., 93 (1960), 99–128. |
| [19] | H. Hermes, Discontinuous vector fields and feedback control, in Differential Equations and Dynamical Systems (eds. J. K. Hale and J. P. LaSalle), Academic Press, (1967), 155–165. |
| [20] | A. F. Filippov, Differential Equations with Discontinuous Righthand Sides, Kluwer Academic Publishers Group, 1988. https://doi.org/10.1007/978-94-015-7793-9 |
| [21] | J. M. Coron, L. Rosier, A relation between continuous time-varying and discontinuous feedback stabilization, J. Math. Syst. Estim. Control, 4 (1994), 67–84. |
| [22] |
F. H. Clarke, Y. S. Ledyaev, R. J. Stern, Asymptotic stability and smooth Lyapunov functions, J. Differ. Equations, 149 (1998), 69–114. https://doi.org/10.1006/jdeq.1998.3476 doi: 10.1006/jdeq.1998.3476
|
| [23] | A. Bacciotti, L. Rosier, Liapunov Functions and Stability in Control Theory, Springer-Verlag, 2005. https://doi.org/10.1007/b139028 |
| [24] | K. A. Bidi, L. Almeida, J. M. Coron, Feedback stabilization for a spatial-dependent sterile insect technique model, unpublished work. |
| [25] |
H. Keller, Non-linear observer design by transformation into a generalized observer canonical form, Int. J. Control, 46 (1987), 1915–1930. https://doi.org/10.1080/00207178708934024 doi: 10.1080/00207178708934024
|
| [26] |
A. Astolfi, L. Praly, Global complete observability and output-to-state stability imply the existence of a globally convergent observer, Math. Control Signals Syst., 18 (2006), 32–65. https://doi.org/10.1007/s00498-005-0161-8 doi: 10.1007/s00498-005-0161-8
|
| [27] |
A. J. Krener, A. Isidori, Linearization by output injection and nonlinear observers, Syst. Control Lett., 3 (1983), 47–52. https://doi.org/10.1016/0167-6911(83)90037-3 doi: 10.1016/0167-6911(83)90037-3
|
| [28] |
P. Bernard, L. Praly, V. Andrieu, Observers for a non-lipschitz triangular form, Automatica, 82 (2017), 301–313. https://doi.org/10.1016/j.automatica.2017.04.054 doi: 10.1016/j.automatica.2017.04.054
|
| [29] | D. Boutat, G. Zheng, Observer Design for Nonlinear Dynamical Systems Differential Geometric Methods, Springer Cham, 2021. https://doi.org/10.1007/978-3-030-73742-9 |
| [30] |
M. Boutayeb, D. Aubry, A strong tracking extended kalman observer for nonlinear discrete-time systems, IEEE Trans. Autom. Control, 44 (1999), 1550–1556. https://doi.org/10.1109/9.780419 doi: 10.1109/9.780419
|
| [31] |
K. Reif, R. Unbehauen, Linearisation along trajectories and the extended Kalman filter, IFAC Proc. Vol., 29 (1996), 3898–3903. https://doi.org/10.1016/S1474-6670(17)58288-9 doi: 10.1016/S1474-6670(17)58288-9
|
| [32] |
A. Krener, The convergence of the extended Kalman filter, Lect. Notes Control Inf. Sci., 286 (2003), 173–182. https://doi.org/10.1007/3-540-36106-5_12 doi: 10.1007/3-540-36106-5_12
|
| [33] | A. Valibeygi, K. Vijayaraghavan, A comparative study of extended Kalman filter and an optimal nonlinear observer for state estimation, in 2017 American Control Conference, (2017), 5211–5216. https://doi.org/10.23919/ACC.2017.7963764 |
| [34] | A. Zemouche, M. Boutayeb, G. I. Bara, Observer design for nonlinear systems: An approach based on the differential mean value theorem, in Proceedings of the 44th IEEE Conference on Decision and Control, (2005), 6353–6358. https://doi.org/10.1109/CDC.2005.1583180 |
| [35] | J. M. Coron, Control and Nonlinearity, American Mathematical Soc., 2007. |




Figures(5) / Tables(1)

Kala Agbo Bidi. Feedback stabilization and observer design for sterile insect technique models[J]. Mathematical Biosciences and Engineering, 2024, 21(6): 6263-6288. doi: 10.3934/mbe.2024274







 DownLoad:
DownLoad: