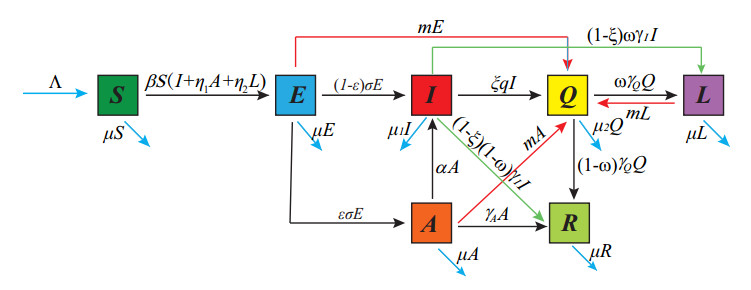
Strangles is one of the most prevalent horse diseases globally. The infected horses may be asymptomatic and can still carry the infectious pathogen after it recovers, which are named asymptomatic infected horses and long-term subclinical carriers, respectively. Based on these horses, this paper establishes a dynamical model to screen, measure, and model the spread of strangles. The basic reproduction number $ \mathcal{R}_0 $ is computed through a next generation matrix method. By constructing Lyapunov functions, we concluded that the disease-free equilibrium is globally asymptotically stable if $ \mathcal{R}_0 < 1 $, and the endemic equilibrium exits uniquely and is globally asymptotically stable if $ \mathcal{R}_0 > 1 $. For example, while studying a strangles outbreak of a horse farm in England in 2012, we computed an $ \mathcal{R}_0 = 0.8416 $ of this outbreak by data fitting. We further conducted a parameter sensitivity analysis of $ \mathcal{R}_0 $ and the final size by numerical simulations. The results show that the asymptomatic horses mainly influence the final size of this outbreak and that long-term carriers are connected to an increased recurrence of strangles. Moreover, in terms of the three control measures implemented to control strangles(i.e., vaccination, implementing screening regularly and isolating symptomatic horses), the result shows that screening is the most effective measurement, followed by vaccination and isolation, which can provide effective guidance for horse management.
Citation: Lusha Shi, Jianghong Hu, Zhen Jin. Dynamics analysis of strangles with asymptomatic infected horses and long-term subclinical carriers[J]. Mathematical Biosciences and Engineering, 2023, 20(10): 18386-18412. doi: 10.3934/mbe.2023817
Strangles is one of the most prevalent horse diseases globally. The infected horses may be asymptomatic and can still carry the infectious pathogen after it recovers, which are named asymptomatic infected horses and long-term subclinical carriers, respectively. Based on these horses, this paper establishes a dynamical model to screen, measure, and model the spread of strangles. The basic reproduction number $ \mathcal{R}_0 $ is computed through a next generation matrix method. By constructing Lyapunov functions, we concluded that the disease-free equilibrium is globally asymptotically stable if $ \mathcal{R}_0 < 1 $, and the endemic equilibrium exits uniquely and is globally asymptotically stable if $ \mathcal{R}_0 > 1 $. For example, while studying a strangles outbreak of a horse farm in England in 2012, we computed an $ \mathcal{R}_0 = 0.8416 $ of this outbreak by data fitting. We further conducted a parameter sensitivity analysis of $ \mathcal{R}_0 $ and the final size by numerical simulations. The results show that the asymptomatic horses mainly influence the final size of this outbreak and that long-term carriers are connected to an increased recurrence of strangles. Moreover, in terms of the three control measures implemented to control strangles(i.e., vaccination, implementing screening regularly and isolating symptomatic horses), the result shows that screening is the most effective measurement, followed by vaccination and isolation, which can provide effective guidance for horse management.

| [1] | H. Qin, M. Zhu, Status of horse breeding in China, Chin. J. Anim. Husb. Vet. Med., (2013), 4–5. |
| [2] | C. R. Sweeney, J. F. Timoney, J. R. Newton, M. T. Hines, Streptococcus equi infections in horses: Guidelines for treatment, control, and prevention of strangles, J. Vet. Intern. Med., 19 (2005), 123–134. |
| [3] | Wellcomecollection, Ruffo, Giordano (d.c.1256), De medicina equorum, 1999. Available from: https://wellcomecollection.org/works/cf2cmy6g. |
| [4] |
J. F. Timoney, Strangles, Vet. Clin. North Am. Equine Pract., 9 (1993), 365–374. https://doi.org/10.1016/s0749-0739(17)30403-0 doi: 10.1016/s0749-0739(17)30403-0

|
| [5] |
N. Pusterla, J. L. Watson, V. K. Affolter, K. G. Magdesian, W. D. Wilson, G. P. Carlson, Purpura haemorrhagica in 53 horses, Vet. Rec., 153 (2003), 118–121. https://doi.org/10.1136/vr.153.4.118 doi: 10.1136/vr.153.4.118
|
| [6] | C. R. Sweeney, R. H. Whitlock, D. A. Meirs, S. C. Whitehead, S. O. Barningham, Complications associated with Streptococcus equi infection on a horse farm, J. Am. Vet. Med. Assoc., 191 (1987), 1446–1448. |
| [7] |
S. E. Heath, R. J. Geor, H. Tabel, K. McIntosh, Unusual patterns of serum antibodies to Streptococcus equi in two horses with purpura hemorrhagica, J. Vet. Intern. Med., 5 (1991), 263–267. https://doi.org/10.1111/j.1939-1676.1991.tb03132.x doi: 10.1111/j.1939-1676.1991.tb03132.x
|
| [8] |
P. A. S. Ivens, D. Matthews, K. Webb, J. R. Newton, K. Steward, A. S. Waller, et al., Molecular characterisation of 'strangles' outbreaks in the UK: the use of M-protein typing of Streptococcus equi ssp. equi, Equine Vet. J., 43 (2011), 359–364. https://doi.org/10.1111/j.2042-3306.2010.00177.x doi: 10.1111/j.2042-3306.2010.00177.x
|
| [9] |
A. S. Waller, New perspectives for the diagnosis, control, treatment, and prevention of strangles in horses, Vet. Clin. North Am. Equine Pract., 30 (2014), 591–607. https://doi.org/10.1016/j.cveq.2014.08.007 doi: 10.1016/j.cveq.2014.08.007
|
| [10] | Ministry of Agriculture and Rural Affairs of the People's Republic of China, Announcement of the Ministry of Agriculture and Rural Development of the People's Republic of China No. 573, 2022. Available from: http://www.moa.gov.cn/govpublic/xmsyj/202206/t20220629_6403635.htm. |
| [11] |
A. G. Boyle, J. F. Timoney, J. R. Newton, M. T. Hines, A. S. Waller, B. R. Buchanan, Streptococcus equi infections in horses: guidelines for treatment, control, and prevention of strangles revised consensus statement, J. Vet. Intern. Med., 32 (2018), 633–647. https://doi.org/10.1111/jvim.15043 doi: 10.1111/jvim.15043
|
| [12] |
S. D. Taylor, W. D. Wilson, Streptococcus equi subsp. equi (Strangles) infection, Clin. Tech. Equine Pract., 5 (2006), 211–217. https://doi.org/10.1053/j.ctep.2006.03.016 doi: 10.1053/j.ctep.2006.03.016
|
| [13] | H. J. Hamlen, J. F. Timoney, R. J. Bell, Epidemiologic and immunologic characteristics of Streptococcus equi infection in foals, J. Am. Vet. Med. Assoc., 204 (1994), 768–775. |
| [14] | A. Boyle, Streptococcus equi subspecies equi infection (strangles) in horses, Compend. Contin. Educ. Vet., 33 (2011), 1–7. |
| [15] |
K. Verheyen, J. R. Newton, N. C. Talbot, M. N. de Brauwere, N. Chanter, Elimination of guttural pouch infection and inflammation in asymptomatic carriers of Streptococcus equi, Equine Vet. J., 32 (2000), 527–532. https://doi.org/10.2746/042516400777584703 doi: 10.2746/042516400777584703
|
| [16] |
J. R. Newton, K. Verheyen, N. C. Talbot, J. F. Timoney, J. L. Wood, K. H. Lakhani, et al., Control of strangles outbreaks by isolation of guttural pouch carriers identified using PCR and culture of Streptococcus equi, Equine Vet. J., 32 (2000), 515–526. https://doi.org/10.2746/042516400777584721 doi: 10.2746/042516400777584721
|
| [17] |
C. Fintl, P. M. Dixon, T. J. Brazil, R. S. Pirie, B. C. McGorum Endoscopic and bacteriological findings in a chronic outbreak of strangles, Vet. Rec., 147 (2000), 480–484. https://doi.org/10.1136/vr.147.17.480 doi: 10.1136/vr.147.17.480
|
| [18] |
A. S. Waller, Strangles: Taking steps towards eradication, Vet. Microbiol., 167 (2013), 50–60. https://doi.org/10.1016/j.vetmic.2013.03.033 doi: 10.1016/j.vetmic.2013.03.033
|
| [19] |
W. O. Kermack, A. G. McKendrick, A contribution to the mathematical theory of epidemics, Proc. R. Soc. A, 115 (1927), 700–721. https://doi.org/10.1098/rspa.1927.0118 doi: 10.1098/rspa.1927.0118
|
| [20] |
O. Byambasuren, M. Cardona, K. Bell, J. Clark, M. McLaws, P. Glasziou, Estimating the extent of asymptomatic COVID-19 and its potential for community transmission: systematic review and meta-analysis, J. Assoc. Med. Microbiol. Infect. Dis. Can., 5 (2020), 223–234. https://doi.org/10.3138/jammi-2020-0030 doi: 10.3138/jammi-2020-0030
|
| [21] |
L. Furuya-Kanamori, M. Cox, G. J. Milinovich, R. J. S. Magalhaes, I. M Mackay, L. Yakob, Heterogeneous and dynamic prevalence of asymptomatic influenza virus infections, Emerg. Infect. Dis., 22 (2016), 1052–1056. https://doi.org/10.3201/eid2206.151080 doi: 10.3201/eid2206.151080
|
| [22] |
N. E. Dean, M. E. Halloran, Y. Yang, I. M. Longini, Transmissibility and pathogenicity of Ebola virus: a systematic review and meta-analysis of household secondary attack rate and asymptomatic infection, Clin. Infect. Dis., 62 (2016), 1277–1286. https://doi.org/10.1093/cid/ciw114 doi: 10.1093/cid/ciw114
|
| [23] |
M. Grunnill, An exploration of the role of asymptomatic infections in the epidemiology of dengue viruses through susceptible, asymptomatic, infected and recovered (SAIR) models, J. Theor. Biol., 439 (2018), 195–204. https://doi.org/10.1016/j.jtbi.2017.12.009 doi: 10.1016/j.jtbi.2017.12.009
|
| [24] |
J. B. Aguilar, J. B. Gutierrez, An epidemiological model of malaria accounting for asymptomatic carriers, Bull. Math. Biol., 82 (2020), 42. https://doi.org/10.1007/s11538-020-00717-y doi: 10.1007/s11538-020-00717-y
|
| [25] |
D. Kalajdzievska, M. Y. Li, Modeling the effects of carriers on transmission dynamics of infectious diseases, Math. Biosci. Eng., 8 (2011), 711–722. https://doi.org/10.3934/mbe.2011.8.711 doi: 10.3934/mbe.2011.8.711
|
| [26] |
L. Matthews, P. G. Coen, J. D. Foster, N. Hunter, M. E. Woolhouse, Population dynamics of a scrapie outbreak, Arch. Virol., 146 (2001), 1173–1186. https://doi.org/10.1007/s007050170113 doi: 10.1007/s007050170113
|
| [27] |
A. Stegeman, A. R. W. Elbers, J. Smak, M. C. M. de Jong, Quantification of the transmission of classical swine fever virus between herds during the 1997–1998 epidemic in The Netherlands, Prev. Vet. Med., 42 (1999), 219–234. https://doi.org/10.1016/s0167-5877(99)00077-x doi: 10.1016/s0167-5877(99)00077-x
|
| [28] |
M. Li, G. Sun, Y. Wu, J. Zhang, Z. Jin, Transmission dynamics of a multi-group brucellosis model with mixed cross infection in public farm, Appl. Math. Comput., 237 (2014), 582–594. https://doi.org/10.1016/j.amc.2014.03.094 doi: 10.1016/j.amc.2014.03.094
|
| [29] | P. J. Hudson, A. P. Rizzoli, B. T. Grenfell, J. A. P. Heesterbeek, A. P. Dobson, The Ecology of Wildlife Diseases, Oxford University Press, Oxford, 2002. |
| [30] |
R. M. A. C. Houben, K. van Maanen, J. G. Kemp-Symonds, A. S. Waller, M. M. S. van Oldruitenborgh-Oosterbaan, H. Heesterbeek, Estimation of the basic reproduction number for Streptococcus equi spp. equi outbreaks by meta-analysis of strangles outbreak reports, Equine Vet. J., 55 (2023), 506–514. https://doi.org/10.1111/evj.13865 doi: 10.1111/evj.13865
|
| [31] |
P. van den Driessche, J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002), 29–48. https://doi.org/10.1016/S0025-5564(02)00108-6 doi: 10.1016/S0025-5564(02)00108-6
|
| [32] |
Z. Shuai, P. van den Driessche, Global stability of infectious disease models using lyapunov functions, SIAM J. Appl. Math., 73 (2013), 1513–1532. https://doi.org/10.1137/120876642 doi: 10.1137/120876642
|
| [33] | J. P. La Salle, The stability of dynamical systems, in CBMS-NSF Regional Conference Series in Applied Mathematics, (1976). |
| [34] | R. A. Horn, C. R. Johnson, Matrix Analysis, 2$^{nd}$ edition, Cambridge University Press, Cambridge, 2012. |
| [35] |
B. Song, Basic reinfection number and backward bifurcation, Math. Biosci. Eng., 18 (2021), 8064–8083. https://doi.org/10.3934/mbe.2021400 doi: 10.3934/mbe.2021400
|
| [36] |
U. Christmann, C. Pink, Lessons learned from a strangles outbreak on a large Standardbred farm, Equine Vet. Educ., 29 (2017), 138–143. https://doi.org/10.1111/eve.12451 doi: 10.1111/eve.12451
|
| [37] |
N. J. Parkinson, C. Robin, J. R. Newton, J. Slater, A. S. Waller, Molecular epidemiology of strangles outbreaks in the UK during 2010, Vet. Rec., 168 (2011), 666. https://doi.org/10.1136/vr.d1485 doi: 10.1136/vr.d1485
|
| [38] |
A. N. F. Neamat-Allah, H. M. E. Damaty, Strangles in Arabian horses in Egypt: Clinical, epidemiological, hematological, and biochemical aspects, Vet. World, 9 (2016), 820–826. https://doi.org/10.14202/vetworld.2016.820-826 doi: 10.14202/vetworld.2016.820-826
|
| [39] |
A. McGlennon, A. Waller, K. Verheyen, J. Slater, J. Grewar, D. Aanensen, et al., Surveillance of strangles in UK horses between 2015 and 2019 based on laboratory detection of Streptococcus equi, Vet. Rec., 189 (2021), 948. https://doi.org/10.1002/vetr.948 doi: 10.1002/vetr.948
|
| [40] |
J. R. Newton, J. L. N. Wood, K. A. Dunn, M. N. DeBrauwere, N. Chanter, Naturally occurring persistent and asymptomatic infection of the guttural pouches of horses with Streptococcus equi, Vet. Rec., 140 (1997), 84–90. https://doi.org/10.1136/vr.140.4.84 doi: 10.1136/vr.140.4.84
|
| [41] |
D. Chhabra, T. Bhatia, U. Goutam, A. Manuja, B. Kumar, Strangles in equines: An overview, Microb. Pathog., 178 (2023), 106070. https://doi.org/10.1016/j.micpath.2023.106070 doi: 10.1016/j.micpath.2023.106070
|











Figures(12) / Tables(2)

Lusha Shi, Jianghong Hu, Zhen Jin. Dynamics analysis of strangles with asymptomatic infected horses and long-term subclinical carriers[J]. Mathematical Biosciences and Engineering, 2023, 20(10): 18386-18412. doi: 10.3934/mbe.2023817







 DownLoad:
DownLoad: