Citation: Jinlong Lv, Songbai Guo, Jing-An Cui, Jianjun Paul Tian. Asymptomatic transmission shifts epidemic dynamics[J]. Mathematical Biosciences and Engineering, 2021, 18(1): 92-111. doi: 10.3934/mbe.2021005

| [1] | E. J. Nelson, J. B. Harris, J. G. Morris, S. B. Calderwood, A. Camilli, Cholera transmission: the host, pathogen and bacteriophage dynamic, Nat. Rev. Microbiol., 7 (2009), 693-702. |
| [2] | C. Lopez, A. C. Dykes, D. D. Sinclair, S. P. Sinclair, J. M. Conn, R. W. Christie, et al., Wateborne giardiasis: a communitywide outbreak of disease and a high rate of asymptomatic infection, Am. J. Epidemiol., 112 (1980), 495-507. |
| [3] | C. Okpalanwa, M. U. Anyanwu, M. A. Momoh, P. O. Nnamani, A. A. Attama, Generic salmonella in asymptomatic adult volunteers: occurrence, antibiogram, extended-spectrum β-lactamase production and carbapenem resistance, Notulae Scientia Biologicae, 11 (2019), 383-390. |
| [4] |
M. A. Montgomery, M. Elimelech, Water and sanitation in developing countries: including Health in the equation, Environ. Sci. Technol., 41 (2007), 17-24. doi: 10.1021/es072435t

|
| [5] | World Health Organization, Drinking-water, available from: https://www.who.int/news-room/factsheets/detail/drinking-water (Accessed 14 June 2019). |
| [6] | W. O. Kermack, A. G. McKendrick, A contribution to the mathematical theory of epidemics, P. Roy. Soc. A-Math. Phy., 115 (1927), 700-721. |
| [7] | V. Capasso, S. L. Paveri-Fontana, A mathematical model for the 1973 cholera epidemic in the european mediterranean region, Revue d'Épidémiologie et de Santé Publique, 27 (1979), 121-132. |
| [8] | C. T. Codeço, Endemic and epidemic dynamics of cholera: the role of the aquatic reservoir, BMC Infect. Dis., 1 (2001), 1. |
| [9] |
J. P. Tian, J. Wang, Global stability for cholera epidemic models, Math. Biosci., 232 (2011), 31-41. doi: 10.1016/j.mbs.2011.04.001
|
| [10] |
J. H. Tien, D. J. D. Earn, Multiple transmission pathways and disease dynamics in a waterborne pathogen model, B. Math. Biol., 72 (2010), 1506-1533. doi: 10.1007/s11538-010-9507-6
|
| [11] |
A. K. Misra, V. Singh, A delay mathematical model for the spread and control of water borne diseases, J. Theor. Biol., 301 (2012), 49-56. doi: 10.1016/j.jtbi.2012.02.006
|
| [12] | M. C. Eisenberg, Z. Shuai, J. H. Tien, P. Van den Driessche, A cholera model in a patchy environment with water and human movement, Math. Biosci., 246 (2013), 105-112. |
| [13] | Y. Wang, J. Cao, Global dynamics of a network epidemic model for waterborne diseases spread, Appl. Math. Comput., 237 (2014), 474-488. |
| [14] | T. Lääveri, Travellers' diarrhoea: aetiological agents revisited, Dissertation, University of Helsinki, Helsinki, Finland, 2019. |
| [15] |
K. M. Angelo, K. A. Jackson, K. K. Wong, R. M. Hoekstra, B. R. Jackson, Assessment of the Incubation Period for Invasive Listeriosis, Clin. Infect. Dis., 63 (2016), 1487-1489. doi: 10.1093/cid/ciw569
|
| [16] | T. Chen, R. Ka-kit Leung, Z. Zhou, R. Liu, X. Zhang, L. Zhang, Investigation of key interventions for shigellosis outbreak control in China, PLoS One, 9 (2014), e95006. |
| [17] | Q. Chen, J. Rui, Q. Hu, Y. Peng, H. Zhang, Z. Zhao, et al., Epidemiological characteristics and transmissibility of shigellosis in Hubei Province, 2005-2017, BMC Infect. Dis., 20 (2020), 272. |
| [18] |
T. Chen, J. Rui, Q. Wang, Z. Zhao, J. Cui, L. Yin, A mathematical model for simulating the phase-based transmissibility of a novel coronavirus, Infect. Dis. Poverty, 9 (2020), 1-8. doi: 10.1186/s40249-019-0617-6
|
| [19] |
P. V. D Driessche, J. Watmough, Reproduction numbers and sub-threshold endemic equilibria forcompartmental models of disease transmission, Math. Biosci., 180 (2002), 29-48. doi: 10.1016/S0025-5564(02)00108-6
|
| [20] |
S. Guo, W. Ma, Remarks on a variant of Lyapunov-LaSalle theorem, Math. Biosci. Eng., 16 (2019), 1056-1066. doi: 10.3934/mbe.2019050
|
| [21] |
J. Arino, F. Brauer, P. van den Driessche, J. Watmough, J. Wu, Simple models for containment of a pandemic, J. R. Soc. Interface, 3 (2006), 453-457. doi: 10.1098/rsif.2006.0112
|
| [22] | National Bureau of Statistics, National data. Available from: http://data.stats.gov.cn/easyquery.htm?cn=C01 (Accessed 28 February 2020). |
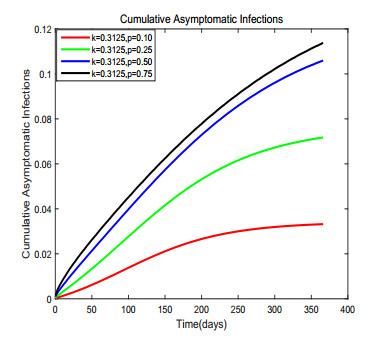
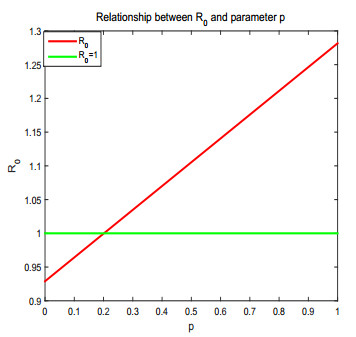
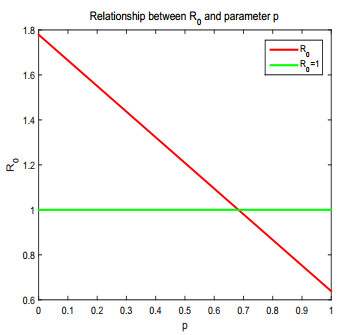
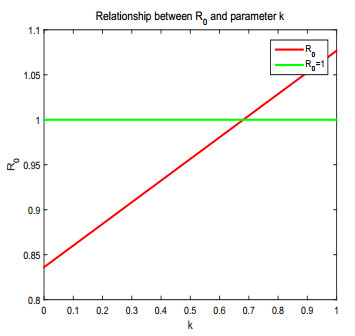
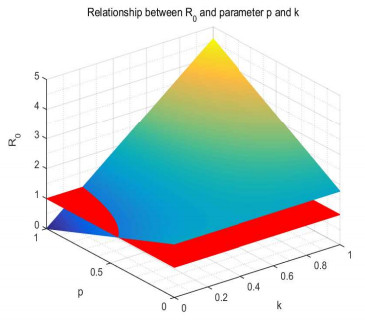
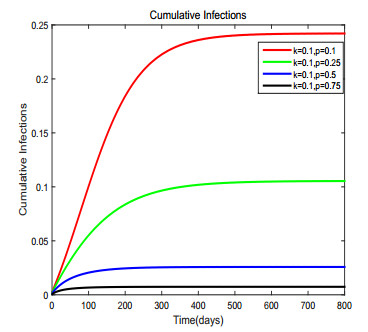
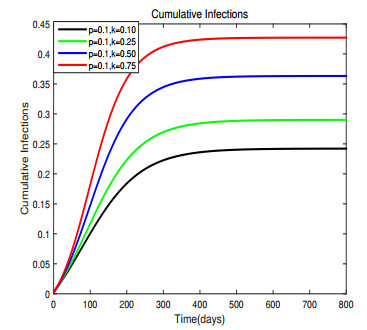
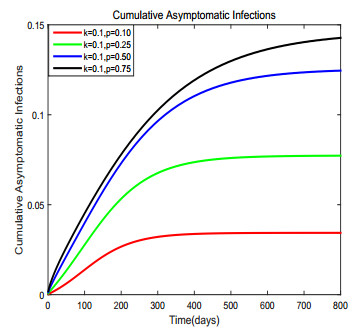
Figures(17) / Tables(1)

Jinlong Lv, Songbai Guo, Jing-An Cui, Jianjun Paul Tian. Asymptomatic transmission shifts epidemic dynamics[J]. Mathematical Biosciences and Engineering, 2021, 18(1): 92-111. doi: 10.3934/mbe.2021005







 DownLoad:
DownLoad: